
1. Introduction
Tilapia (Oreochromis niloticus) is recognized as a fast-growing farmed fish species with simple husbandry techniques. The fish species is one of the most popular aquaculture commodities in different parts of the world, providing animal protein for all levels of societies (Amal et al. 2018; Suhermanto et al. 2019; T. Taukhid et al. 2023). According to FAO (2017), after the Cyprinidae family tilapia is the second largest aquaculture commodity globally. It accounts for about 5% of global aquaculture production. China, Indonesia, and Egypt are the top three tilapia producers worldwide. In 2019, Indonesia produced 1,506,156 tons of tilapia valued at IDR 34.7 trillion (MMAF, 2021).
Over the past decade, a novel disease is affecting tilapia. The disease named Tilapia Lake Virus Disease (TiLVD), has emerged, caused by a newly discovered viral genus within the family Orthomyxoviridae (Eyngor et al. 2014). Subsequent genetic analysis reclassified the pathogen to the Tilapinevirus genus and Amnoonviridae family (Adams et al. 2017). TiLV disease was initially known to affect only tilapia (Dong et al. 2017; Eyngor et al. 2014; Fathi et al. 2017; Surachetpong, Roy, and Nicholson 2020), however, several fish species are currently known to be susceptible to the virus (Abdullah et al. 2018; Bergmann et al. 2021; Jaemwimol et al. 2018; T. Taukhid et al. 2023; Yamkasem et al. 2021). During the TiLV outbreak, differences in mortality rates were noted among different tilapia strains (Ferguson et al. 2014; Tattiyapong, Dachavichitlead, and Surachetpong 2017).
The spread of TiLV has spread to over 15 countries across four continents, including Indonesia (Aich et al. 2022; DGA 2017; Koesharyani et al. 2018; Surachetpong, Roy, and Nicholson 2020). Despite more than 25 tilapia strains being farmed in Indonesia, none have been specifically identified as resistant to TiLV. Nonetheless, variances in resistance to Streptococcus agalactiae bacteria have been documented among Indonesian tilapia strains such as Best, Gesit, Nirwana, Srikandi, Gift, Aureus, Sultana, and Red Tilapia (Febrianti, Taukhid, and Lusiastuti 2015; E. A. Taukhid 2014). Similar genetic variations associated with resistance to bacterial infections have been observed in tilapia strains elsewhere (LaFrentz et al. 2016; Wonmongkol et al. 2018).
This study aims to evaluate the resistance of six Indonesian tilapia strains to TiLV disease and assess the impact of mimicked daily water temperature fluctuations on the disease severity. The findings contribute technical insights, including strain selection based on regional TiLVD prevalence and temperature control measures, to mitigate the adverse effects of TiLV-related disease outbreaks.
2. Materials and Methods
2.1. Fish
Six tilapia strains (Table 1) used in this study were obtained from the Ministry of Marine Affairs and Fisheries (MMAF) or local farms, sourced either from primary breeders or certified hatcheries with traceable origins. Prior to introduction into the facilities, a sub-sample comprising 20 fish from each tilapia strain (n=120 fish) were tested for the presence of TiLV, Streptococcus agalactiae, and Aeromonas hydrophila. The presence of TiLV was performed on specific target organs (Eyngor et al. 2014) and analysed by semi-nested RT-PCR using two primer sets. The cycling conditions and reagents used were consistent with those described by Dong et al. (2017). Bacteria were isolated from kidney, spleen and liver. Bacteria isolated from each organ were grown separately on tryptic soy agar (DifcoTM) and/or Brain Heart Infusion Agar (DifcoTM). Bacterial analysis included Gram stain, motility, oxidase, Rimmler-Shott, catalase, O/F test, triple sugar iron agar (TSIA) and other biochemical reactions using API 20 E and API 20 Strep (Biomereux). Identification was performed according to SNI 7545.3 (2009); SNI 7303.1 (2015); Austin and Austin (2016). Fish were acclimated for three weeks in individual concrete recirculating tanks, each measuring 2 x 4 x 0.8 m3 and containing 500 fish of a single strain. They were fed commercial pellets diet (protein≈28%), at 5% of the fish’s body weight per day dispensed twice daily. During the acclimation period, water quality parameters were closely monitored and maintained at stable levels, DO levels above 4.0 ppm, water temperature 27 - 29°C, and pH levels 6.8 to 7.0.
2.2. Source of infective TiLV
A population of TiLV (+) tilapia collected from an outbreak in Bogor area was used as a source of experimental infection. To maintain the source of infection, 200 infected fish from the natural TiLV outbreak in Bogor were placed in individual 500-litre circular fibreglass tanks with aeration and filters (tank 1), while approximately 400 non-specific tilapia free of TiLV infection were placed in another tank (tank 2). Every day for three weeks, 10 healthy fish were added to the TiLV tank, and the number of fish that died was also replaced. Every 2 days, organs from 5 fish from tank 1 were pooled and analysed. were tested by semi-nested RT-PCR (Dong et al. 2017) to check TILV status.
2.3. Experimental design
TiLV disease resistance was tested in a controlled system using 100 litre plastic tanks filled with 80 litres of water. Two infection methods, by cohabitation and intraperitoneal (IP) injection were used.
Cohabitation experiments, two distinct experimental environments were applied. The first was a closed-system with regulated water temperature ranging from 30.0 to 32.5°C and water recirculation. The second was a semi-closed system lacking water recirculation and temperature regulation but with aeration and individual covers.
In the cohabitation method, each tank was stocked with 20 test fish and 5 TiLV (+) fish in three replicates. Fish from the infected-source (infective TiLV) were identified by cutting a small part of the pelvic fin. If any of these infected fish died during the study, they were replaced with new infected-source fish to maintain TiLV contamination. Negative control groups for each strain were kept separately, but the overall system was the same as the treatment groups.
Intraperitoneal injection, the procedure was performed in a closed system with regulated water temperature ranging from 30.0 to 32.5°C and water recirculation. As a specific cell culture for TiLV was not available in our laboratory, we did not use the standard tissue culture infection dose (TCID) to determine the concentration of inoculated virus. Instead, in this study, we assumed that the concentration of virus administered to the fish remained consistent, due to viral homogenisation procedure outlined as follows: (a). The target organs (liver, spleen, brain, eyes, gills, and kidneys) from 25 tilapia TiLV-positive (+) were aseptically removed, homogenised, diluted with Minimum Essential Medium (MEM) 1:9 (w/v), and centrifuged at 10,000 rpm for 5 min. (b). The supernatant was filtered through a 0.22 µm Millipore filter to obtain TiLV virus homogenate solution, and (c). The quality of the homogenate was tested for the presence of TiLV virus by semi-nested RT-PCR and grown on bacterial media to ensure that only the virus was present in the solution.
A total of 60 fish from each tilapia strain were artificially infected with TiLV homogenate at a rate of 0.1 mL per fish, divided into triplicates. Before injection, the fish were anesthetised with a commercial anaesthetic agent (Nishikigoi Stabilizer Anaesthetic). The negative control group received an injection of phosphate-buffered saline (PBS) and was kept separately, while the overall system remained the same as the treatment groups.
2.4. Laboratory and statistical analysis
Behaviour, clinical signs, fish mortality, and water temperature were observed daily over a 24-day period. On day 7 post-infection, three randomly selected fish from each treatment group were sampled for TiLV detection. For TiLV detection, selected organs (eyes, brain, liver, spleen, and kidney) were pooled and preserved in RNA later solution and/or a solution containing 80:20 (v/v) absolute ethanol:glycerol, then stored at -70°C until analysis. A portion of these organs were pooled until a total weight of 1 gram, and then subjected to extraction for further testing.
Semi-nested RT-PCR was performed using two sets of primers: ext-1 (TATGCAGTACTTTCCCTGCC) and ME1 (GTTGGGCACAAGGCATCCTA) in step-1 nested RT-PCR with a target PCR amplicon of 415 bp. Step-2 Semi-nested with primers: ME2 (TATCACGTGCGTACTCGTTCAGT) and ME1 (GTTGGGCACAAGGCATCCTA) with a target PCR band of 250 bp. All of the cycling conditions and reagents were identical to those used by Dong et al. (2017).
Bacterial isolation was performed on kidney, spleen, and liver samples, which were aseptically plated on Tryptic Soy Agar (DifcoTM) and/or Brain Heart Infusion Agar (DifcoTM). Bacterial analysis included Gram stain, motility, oxidase, Rimmler-Shott, catalase, O/F test, Triple Sugar Iron Agar (TSIA), and other biochemical reactions using API 20 E and API 20 Strep (Biomerieux). Identification was carried out according to SNI 7545.3 (2009) and SNI 7303.1, as well as procedures described by Austin and Austin (2016).
Statistical differences in the resistance of tilapia strains to TiLV disease were assessed using Anova analysis of variance, with differences between tilapia strains analysed by least significant difference (LSD). Differences in percentage mortality between temperature-controlled and uncontrolled temperature conditions were analysed using z-test analysis. Significant differences were determined at α = 0.05. The relative percentage survival (RPS) of genetically improved strains compared to unimproved strains was calculated using the following formula:
RPS = [ (survival in genetically improved strain /survival in non-improved strain) -1] x 100
3. Results
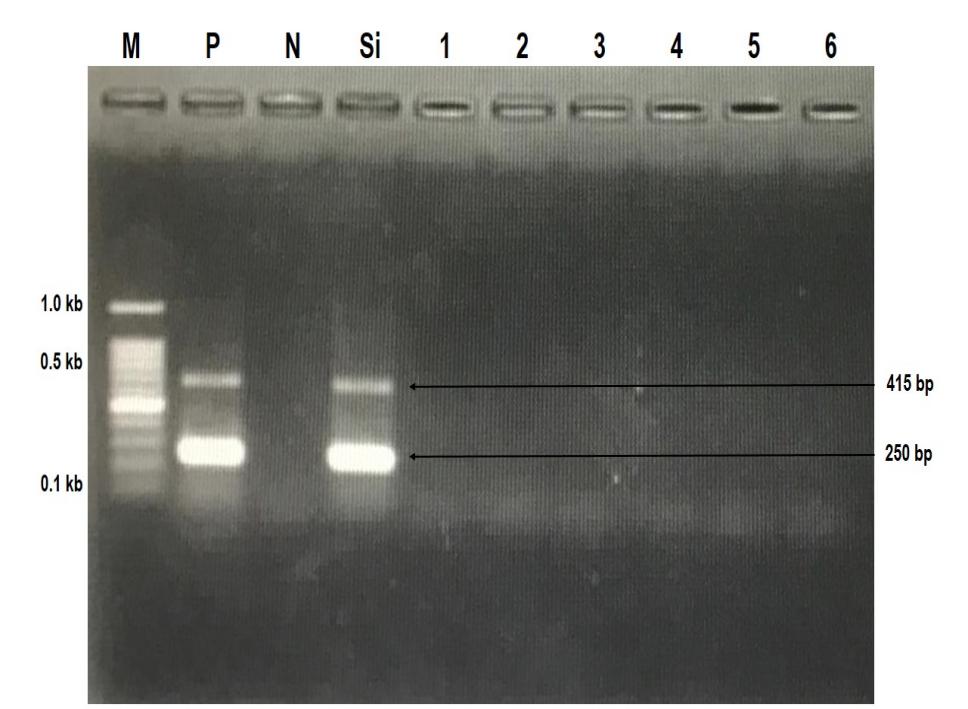
During the acclimation period, random samples were tested for the presence of TiLV, and the results were negative for the virus, as shown in Figure 1, and were free of concomitant bacterial pathogens (S. agalactiae and A. hydrophila). These results suggest that the tilapia strains used in this study can be considered as specific pathogen-free (SPF) for TiLV disease.
_detected_in_six_tilapia_strains_p.jpeg)
During the study, a range of changes have been observed noted, including weakness, reduced appetite, darkened body color, dropsy, cataracts, endo/exophthalmus, and erosion of fins/skin. These symptoms are not pathognomonic of TLVD. However, a visually consistent symptom is the change in the golden ring around the eye due to uveitis, and in severe cases, the uvea may not even be visible. A typical description of these clinical signs is shown in Figure 2.
.png)
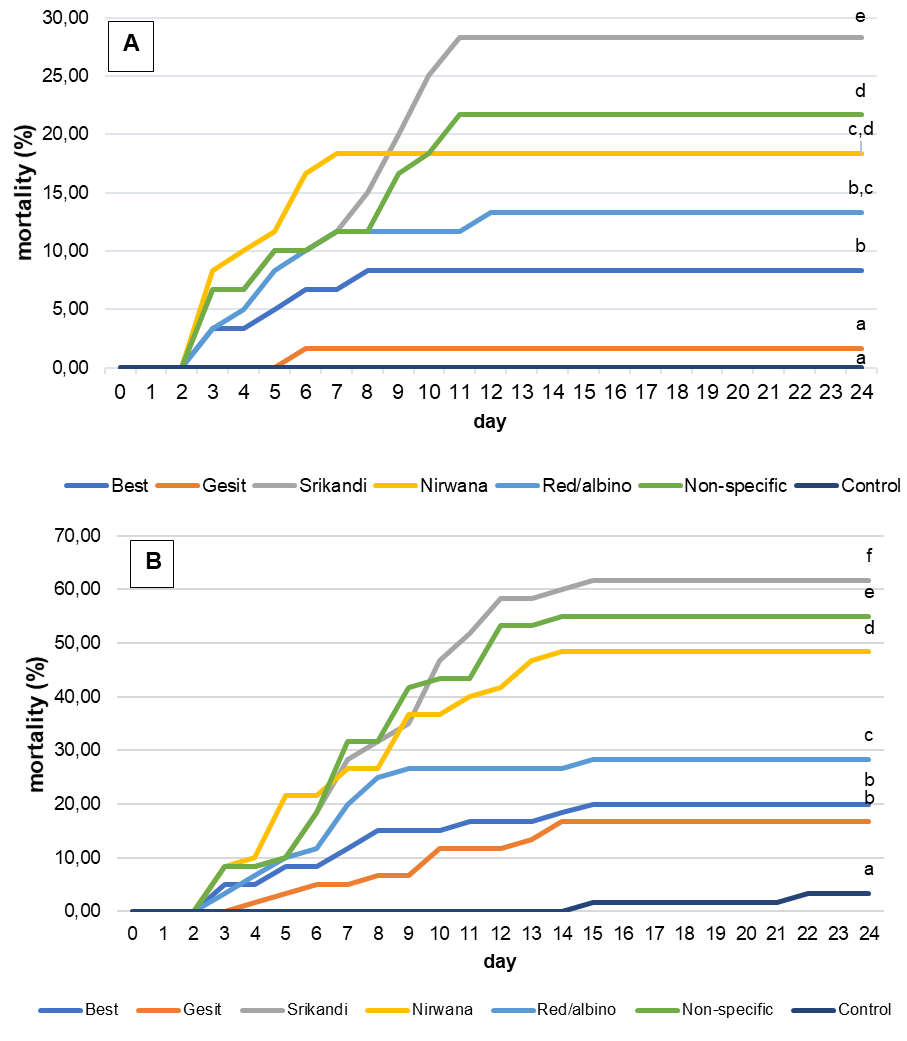
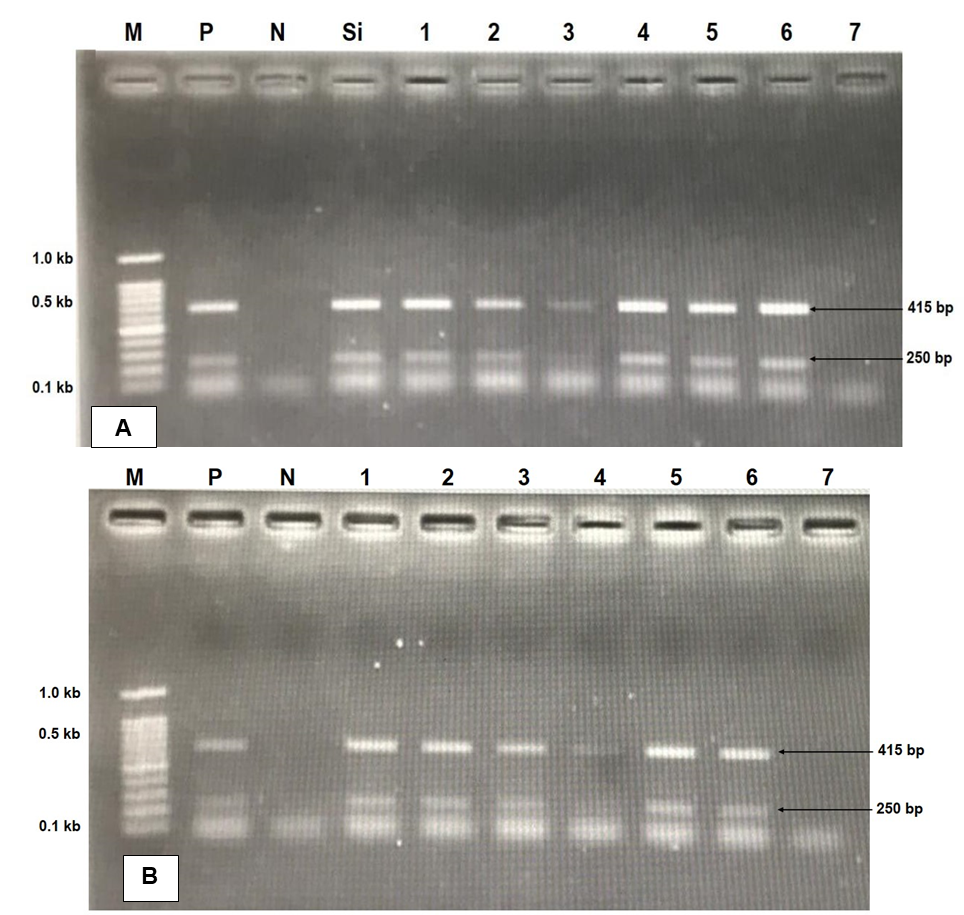
No significant differences were observed between replicates, so mortality data from the same strain were pooled. Mortality occurred mainly between days 4 and 10 post-infection, after which mortality ceased and affected fish entered a healing phase. Tilapia infected by both intraperitoneal and cohabitation methods showed a similar mortality pattern, with mortality peaking between days 3 and 10 (figure 3 A & B). However, fish infected by IP had a higher mortality rate and a prolonged mortality. Significant differences in cumulative mortality were observed between strains, with both infection methods showing comparable patterns. The Gesit strain showed the highest resistance to TiLV disease, with a cumulative mortality of 1.67% for cohabitation (16.70% for IP injection). Conversely, the Srikandi strain had the highest mortality rate, with 28.33% cumulative mortality for cohabitation (61.67% for IP injection). On day 7 after artificial infection, molecular analysis of randomly selected samples from each treatment group (strain) revealed that all six tilapia strains tested positive for TiLV (+), whereas the negative control groups remained negative for TiLV (-) (Fig. 4).
_mortality_rate_and_cumulative_mortality_of_six_strains_of_tilapia_artificially_infected_.png)
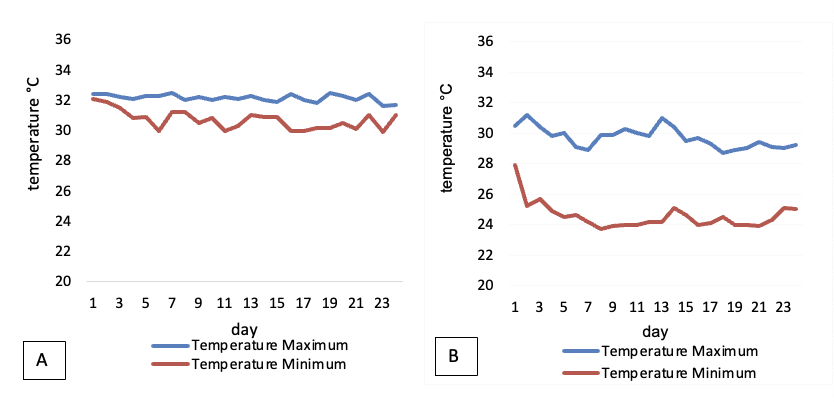
Significant differences in water temperature were observed between the controlled and uncontrolled systems during the experimental cohabitation infection. The controlled system maintained a relatively stable water temperature range of 29.9 - 32.5°C, with the largest fluctuation over a 24-hour period being approximately 2.3°C. In contrast, the uncontrolled system exhibited greater temperature fluctuations, ranging from 23.7 - 31.2°C, with the most significant fluctuation over a 24-hour period being approximately 6.3°C. This is illustrated in Figure 5.
._gel_electrophoresis_profile_of_tilapia_lake_virus_(tilv)_detection_at_7th_days_after_a.png)

The study revealed a remarkable relationship between uncontrolled water temperature and the severity of infections in cohabiting fish. This correlation was evident in all strains except the Best strain, as supported by statistically significant differences observed between the controlled and uncontrolled groups (z-test; Table 2).
At the end of the 24-day observation period, all surviving fish, including those exposed to both IP infection and cohabitation methods, were pooled and placed in a controlled concrete pond. No mortalities were recorded for 30 days after pooling (54 days post-infection). To assess the persistence of TiLV in previously exposed fish, a random sample of 15 individuals was tested, revealing a prevalence of only 1 TiLV-positive case (6.67%), while the vast majority (93.33%) tested negative for TiLV.
Discussion
During TiLV outbreaks, variations in mortality rates were observed among different tilapia strains (Ferguson et al. 2014; Tattiyapong, Dachavichitlead, and Surachetpong 2017), and multiple outbreak reports in Indonesia indicated discrepancies in environmental factors, notably fluctuations in water temperature. These empirical findings imply potential disparities in the susceptibility of tilapia strains to TiLV. Therefore, this study aimed to evaluate the resistance of six Indonesian tilapia strains to TiLV disease and to assess the effects of simulated daily water temperature fluctuations on disease severity.
TiLV disease is known to be transmitted both horizontally and vertically (Eyngor et al. 2014; Ferguson et al. 2014; Liamnimitr et al. 2018). Two artificial transmission models have been investigated: cohabitation and intraperitoneal (IP) injection. Cohabitation more closely mimics natural conditions, but may be subject to various confounding factors, including fish stress levels (Ferguson et al. 2014). Conversely, IP injection provides a scalable artificial infection model commonly used to study pathogenicity and host-pathogen interactions. This study demonstrates that both artificial infection models function effectively, as molecular evidence of viral presence was observed in selected organs on day 7 post-artificial infection.
Consistent with previous findings by Liamnimitr et al. (2018), both artificial infection models exhibited a mortality pattern characterised by an onset on day 3 and a peak on days 4-10. Subsequently, the majority of surviving fish exhibited full recovery by week 3 (Barría et al. 2020; Eyngor et al. 2014). However, at 54 days post-infection, 6.67% (1/15) of apparently healthy samples still tested positive for TiLV. The study provides evidence that genetic origin plays a critical role in influencing resistance to tilapia lake virus disease (TiLVD) in six different tilapia strains. Remarkable differences in cumulative mortality rates were observed among these strains, with both infection methods showing similar patterns.
The genetic characteristics of Indonesian tilapia strains are likely associated with variation in disease resistance (Febrianti, Taukhid, and Lusiastuti 2015; E. A. Taukhid 2014). Furthermore, E. A. Taukhid (2014) identified a linear correlation between the susceptibility of tilapia strains and their genetic composition. Specifically, the tilapia strain susceptible to S. agalactiae harbours specific allele fragments at specific loci that are genetically associated with disease susceptibility. Variation in resistance of tilapia strains to bacterial diseases has been empirically demonstrated (Febrianti, Taukhid, and Lusiastuti 2015; E. A. Taukhid 2014; Wonmongkol et al. 2018).
There are few reports illustrating genetic resistance of TiLV infection, which was strongly suspected based on the data/information available at the time of the disease outbreak (Ferguson et al. 2014; Tattiyapong, Dachavichitlead, and Surachetpong 2017). Tattiyapong, Dachavichitlead, and Surachetpong (2017) demonstrated differential responses to artificial infection of two tilapia strains (red and black) with TiLV, suggesting potential strain-specific susceptibility. Consequently, an effective disease control strategy may involve the development or use of tilapia strains with increased resistance to TiLV (Aich et al. 2022; Barría et al. 2020; OIE (World Organisation for Animal Health) 2017; Tang et al. 2021). Our hypothesis is that survivors can develop targeted immunity to a level of protection that exceeds virus development and pathogenicity. When the virus enters the body, fish activate their innate immune system, followed by antibody production. Genes involved in the interferon pathway include interferon (Yu et al. 2010), myxovirus resistant protein (mx) (Leong et al. 1998), interferon-stimulated gene (Das et al. 2017) and viperin (Wang et al. 2019). The mx gene was detected at significant levels in TiLV-resistant tilapia strains, resulting in significantly reduced TiLV viral load. In addition, the resistant tilapia strains showed low levels of pro-inflammatory response (Adamek et al. 2022).
The study showed that the virulence and impact of TiLV disease is significantly affected by temperature fluctuations. Sustained water temperature fluctuations outside the tolerance threshold would significantly affect the physiology and health status of the fish, making them more susceptible to disease. In tilapia exposed to TiLV, when water temperature fluctuations were < 2°C over 24 hours and sustained, the virulence of TiLV and the impact of the disease were relatively low. However, if the fluctuation is > 4°C over 24 hours and sustained, the virulence and impact of TiLV disease will be significant.
A presumptive diagnosis of TiLV infection in tilapia can be made by observing behavioural changes, clinical signs and other disease-related signs as described in the existing literature (Dong et al., 2017; Jaemwimol et al. 2018; Surachetpong et al. 2017; Tsofack et al. 2017). These changes include weakness, body darkening, dropsy, cataracts, loss of the golden ring around the eye (uvea), and swelling of internal organs (Bacharach et al. 2016; Jaemwimol et al. 2018). However, these signs are also common in tilapia infected with other diseases and cannot be considered specific to TiLV. Although uveal changes are a clear clinical sign, they should not be relied upon alone to diagnose TiLV, as they may not be visible in severe cases, however, molecular detection methods should be used to confirm the diagnosis.
Healthy carriers of TiLV may pose a significant challenge to disease control. Our results indicate that infected tilapia can be healthy carriers almost two months after infection, but only one out of fifteen individuals was found to be positive, suggesting that the persistence of TiLV in fish is influenced by a wide range of internal and external factors. In epidemiological models, the proportion of infectious tilapia appears to be maximal after 5 days of contamination, decreasing but not disappearing after 16 days (Yang et al. 2018), and fish without clinical signs also tested positive for TiLV (Senapin et al. 2018). Considering the survival of the virus outside the host, water disinfection for 3-5 days before stocking new tilapia (Yamkasem et al. 2022) could be an effective biosecurity measure, however, considering the genetic resistance shown in our study, in farming areas where the risk of TiLVD is high, the selected resistant strain could be an efficient preventive measure.
Acknowledgments
This work was supported by the Quick Response Projects of the Agency for Marine and Fisheries Research and Human Resources, Ministry of Marine Affairs and Fisheries. The authors thank Edy Farid W., Ahmad Wahyudi and Setiadi for their technical assistance during the experiments. This is an ISEM publication ISEM 2024-103 SUD
Conflict of interest
Authors declare no conflicts of interest
Data availability statement
The data sets supporting the results and any additional information are available on request, please send an e-mail to tauk001@brin.go.id.
Ethic statement
Fish were treated according to Indonesian regulations, and all experiments were carried out under the accreditation SNI 6141:2009 and SNI 8124:2015.