
Introduction
Motile Aeromonas septicemia (MAS) is one of the most important bacterial diseases in aquaculture caused by various Aeromonas species and results in severe economic losses (Elgendy, Moustafa, et al. 2015). Aeromonads are ubiquitous in aquatic environment, but under stressful conditions, they can cause serious infections and high levels mortality in farmed fish (El-Gohary, Zahran, and Abd El-Gawad 2020). Aeromonas hydrophila, A. sobria, A. caviae, and A. veronii are the most pathogenic Aeromonas spp. in aquaculture (Abdelsalam et al. 2021).
The pathogenesis of MAS is multifactorial and requires the interaction of several variables for the disease to develop. Aeromonas species secrete various extracellular enzymes and possess numerous virulence genes that enable bacteria to overcome the host’s immune system and initiate severe infections (Hu et al. 2012; El-Gohary, Zahran, and Abd El-Gawad 2020).
Nile tilapia (Oreochromis niloticus) aquaculture constitutes an integral part of the Egyptian aquaculture industry (Elgendy et al. 2022). Farmed tilapia are susceptible to numerous bacterial diseases causing massive mortalities and colossal economic losses (Abdelsalam et al. 2022). The excessive use of antibiotics by fish farmers to control such infections have detrimental impacts on the aquatic ecosystem and poses a threat to human health due to the emergence of resistant strains (Elgendy, Shaalan, et al. 2021; Ali et al. 2022; Algammal et al. 2022). This study was conducted to analyze Aeromonas spp. isolates recovered from farmed O. niloticus that suffered massive mortalities during summer. The molecular characteristics, virulence profile, antibiotic resistance and pathogenicity of aeromonads isolates were investigated.
Materials and methods
Case history and sampling
Abnormal mortalities (250 dead fish every day/earthen pond) were recorded in O. niloticus (110–200 g) within five earthen ponds in a fish farm in Beheira Governorate, Egypt, in June 2020. Fish were stocked at a density of 30,000 fish/acre. The farm owner used untreated poultry droppings to promote natural food production in the ponds and had rear-Pecking ducks (190 ducks per/acre). Cattle and sheep grazed at the banks between the earthen ponds. There were several dogs straying on the farm. Moribund fish exhibited signs of hemorrhagic septicemia and respiratory distress. The average temperature, dissolved oxygen level, and unionized ammonia level were 27°C, 4 mg/L, and 0.53 mg/L, respectively. A total of 70 moribund fish were collected and transferred in an ice box to the hydrobiology department laboratory NRC. Water samples (n = 15) (3 samples from each affected pond) were collected and analyzed according to Newaj-Fyzul et al. (2008).
Bacteriological examination
Bacteriological smears were obtained from fish hematopoietic organs (kidneys and liver) and subcultured onto tryptic soy agar. Water samples were collected and examined according to Newaj-Fyzul et al. (2008). Bacterial strains were purified and identified using the Vitek 2 compact device (bio-Merieux).
Molecular identification of Aeromonas spp.
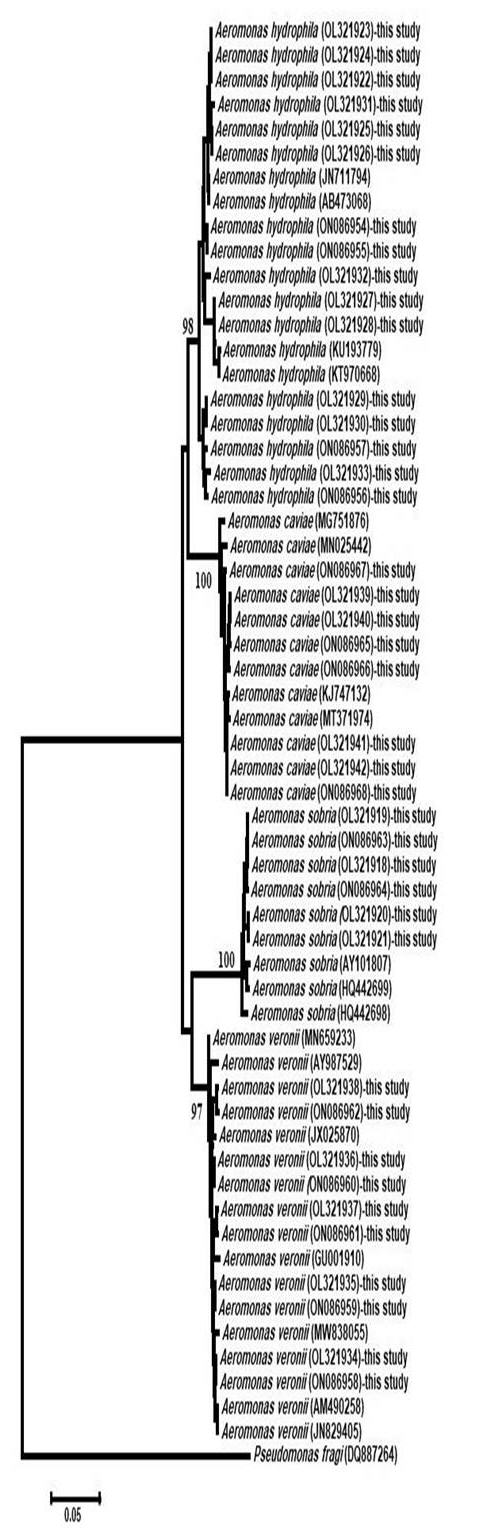
DNA was extracted from purified bacterial strains according to the PrepMan® Ultra Sample Preparation Reagent protocol. Polymerase chain reaction was conducted using gyrB-specific primers as described by Hu et al. (2012). The amplified gyrB was directly sequenced in two directions using the Sanger DNA sequencer ABI 3730xl DNA sequencer (Applied Biosystems™, USA) at Sigma (Cairo, Egypt), assembled using Bio Edit version 7.0 (Hall 1999) and aligned to other interrelated sequences in the GenBank database using BLASTN search (National Center for Biotechnology Information). The neighbor-joining phylogenetic tree was constructed using MEGA version X (Kumar et al. 2018).
Virulence characteristics of Aeromonas spp.
Extracellular enzymes
Lipase and protease activities were analyzed according to Carrazco-Palafox et al. (2018) and Sokol, Ohman, and Iglewski (1979).
Detection of virulence and antibiotic resistance genes
The presence of aerolysin (Aer), hemolysin (Hyl), lateral flagella A (laf-A), elastase (ela), serine protease (Ser), cytotoxic enterotoxin (Act), and glycerophospholipid-cholesterol acyltransferase (gcaT) genes in the recovered aeromonads strains was investigated according to Sun et al. (2016) using specific primers (Table 1). The strains were also analyzed for the presence of the following antibiotics resistance genes: aminoglycosides (aadA), β-lactams (blaTEM), tetracycline (tetC & tetA), and sulphonamide (sul 1) according to Ndi & Barton (2011) using specific primers (Table 1).
Antibiotic susceptibility testing
The susceptibility of the Aeromonas spp. isolates to ampicillin 10 µg, amoxicillin 30 μg, gentamicin 10 μg, trimethoprim/sulfamethoxazole 1.25/23.75 μg, florfenicol 30 µg, ciprofloxacin 5 μg, and tetracycline 30 μg was evaluated using the disk diffusion method according to CLSI (2010).
Pathogenicity
A total of 720 healthy Nile tilapia with an average weight of 55 g were collected and acclimatized in aerated aquaria (50 L each) at 25°C ± 1°C. The fish were divided into groups of 10 fish/aquarium in duplicates. They were anesthetized using MS-222 (Sigma) (150 μgL-1). One isolate representative for each Aeromonas spp. was chosen at random and used to determine the LD50 value. Bacterial cultures were serially diluted, and each fish group was injected intraperitoneally (I/P) with 0.1 ml of the relevant bacterial culture suspension at concentrations ranging from 102 to 109 CFU/ml/fish (one isolate and one challenge dose per tank). Control fish were injected with 0.1 ml of sterile phosphate buffered saline (PBS). All fish were monitored for 13 days. The LD50 value was calculated according to Reed J. and Muencha H. (1938).
Histopathology
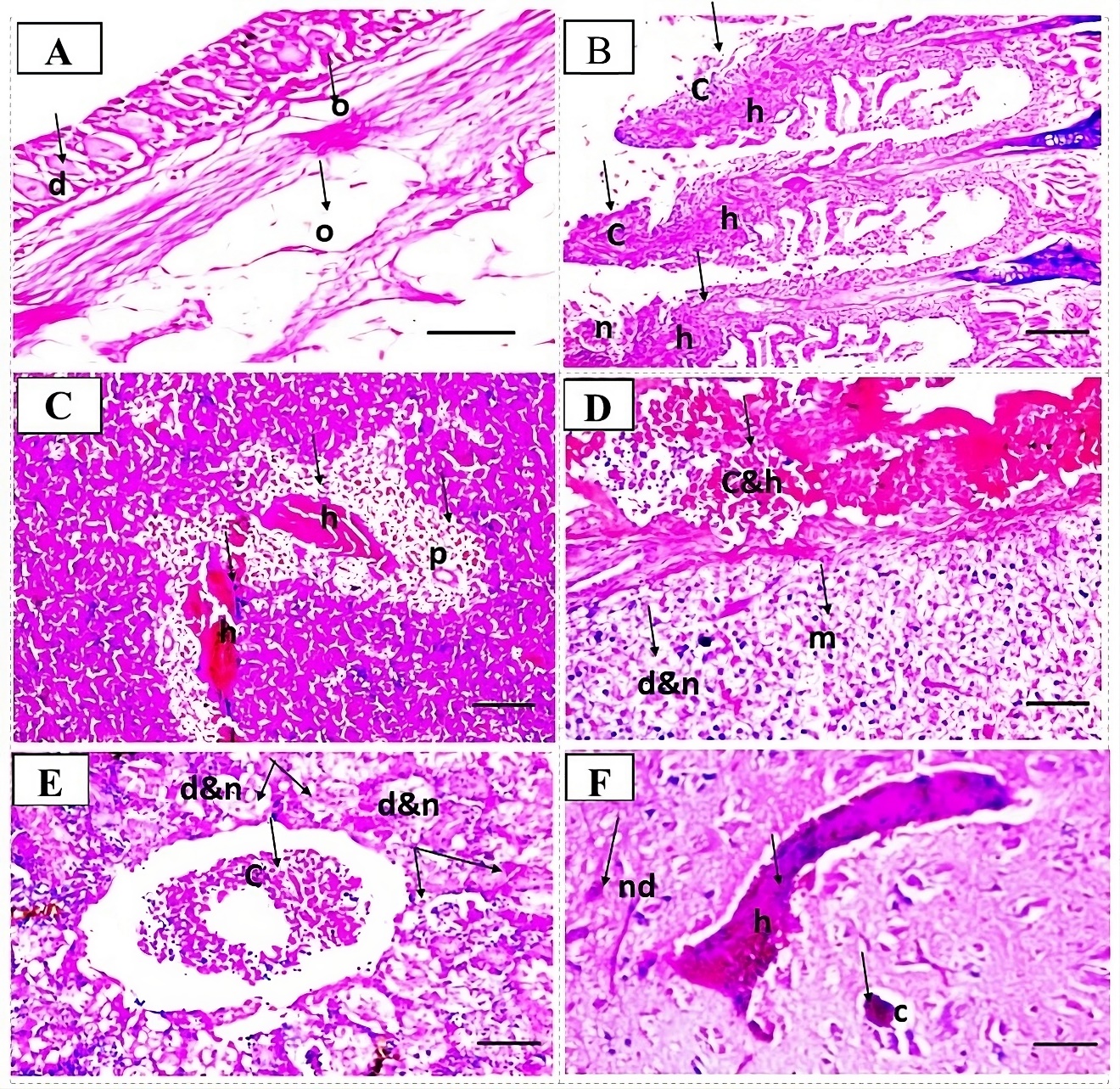
Haematoxylin and eosin (H&E) stained histopathological sections were prepared from tissue specimens collected from experimentally infected fish following the methods described by Bancroft and Gamble (2008).
Results
Clinical examination
Moribund tilapia demonstrated signs of hemorrhagic septicemia. Petechial hemorrhages, fin rot, and skin erosions were widely distributed on the external body surface. Internally, the liver, spleen, and kidney were congested and enlarged. Ascites and exophthalmia were noted in some fish (Fig. 1).
_naturally_infected_tilapia_showing_hemorrhages_on_the_external_body_surface_and_at_the_b.jpeg)
Bacteriological examination
In total, 31 Aeromonas spp. isolates were recovered from the investigated fish specimens and identified as A. hydrophila (n = 12), A. veronii (n = 10), A. caviae (n = 5), and A. sobria (n = 4). Nine Aeromonas spp. isolates were discovered in the farm water samples, including A. hydrophila (n = 4), A. sobria (n = 2), and A. caviae (n = 3). A. veronii was not detected in the water samples. Some nonpathogenic bacteria (Proteus spp., Citrobacter freundii, and Shigella spp.) were also detected in some water and fish samples.
Molecular identification of Aeromonas spp.
A comparison of gyrB sequences revealed that all isolates (n = 40) belonged to the genus Aeromonas, the identity of which was confirmed as A. hydrophila (n = 16), A. caviae (n = 8), A. veronii (n = 10), and A. sobria (n = 6).
A. hydrophila isolated from water samples (OL321924, OL321926, OL321928, and OL321930) and that isolated from fish specimens (OL321922, OL321923, OL321925, OL321927, OL321929, OL321931–OL321933, and ON086954–ON086957) exhibited 99.72%–98.05% similarity to the following A. hydrophila strains: JN711793T, JN711794T, JN711795T, AB436660T, and JN711793T (typing strains) and HQ701864.1, GQ471011.1, JQ805070.1, AB473095.1, KF873661.1, MK484180.1, JN711794.1, and AB473068.1. The intraspecies similarity was 97.74%–100% for 12 isolates of A. hydrophila recovered from tilapia and water, with nucleotide differences ranging from 10 to 24 bp.
A. caviae recovered from fish (OL321939, OL321941, and ON086965–ON086967) and that recovered from water (OL321940, OL321942, and ON086968) showed 98.92%–98.22% similarity to the following A. caviae strains: AJ868400T, JN829530T, and KC924126T (typing strains) and MT371974.1, LC003106.1, MN855498.1, KJ747132.1, MN855516.1, KR140073.1, KC924174.1, and MN855511.1. The intraspecies similarity was 99.19%–100% for four A. caviae isolates recovered from fish and water, with eight nucleotide differences.
The sequence alignment of A. sobria isolated from fish (OL321918, OL321920, ON086963, and ON086964) and that recovered from water (OL321919 and OL321921) exhibited 98.91–98.15% similarity to the following A. sobria strains: AB473084T, HQ442698T, AF417631T (typing strains) and MG263589.1, AB473086.1, JN829516.1, MG263541.1, KP115770.1, KJ743530.1, AY101781.1, and HQ442698.1. The intraspecies similarity was 99.20%–100% for four A. sobria isolates recovered from Nile tilapia and water, with eight nucleotide differences.
The sequence alignment of A. veronii isolated from fish (OL321934–OL321938 and ON086958–ON086962) showed 99.34%–98.84% similarity to the following A. veronii strains: FN796748T, HM584508T, and AF417626T (typing strains) and MN659233.1, LC644255.1, AB829112.1, JF938686.1, KR140071.1, MN025464.1, LC003119.1, and JX025899.1. The intraspecies similarity was 98.56%–100% for five A. veronii isolates recovered from tilapia, with nucleotide differences ranging from 6 to 16 bp. The phylogenetic analysis confirmed the identity of the isolates (Fig. 2).

Virulence characteristics of Aeromonas spp.
Proteolytic and lipolytic activities
Proteolytic and lipolytic activities were noted in (52.5%) and (47.5%) of isolates recovered from fish, respectively, and in (20%) and (17.5%) of isolates from water, respectively.
Detection of virulence and antibiotic resistance genes
Analysis of Aeromonas spp. isolates recovered from fish showed that Ser was the most commonly detected gene (62.5%), followed by Aer (55%), ela (37.5%), gcaT (32.5%), Hyl (25%), laf-A (22.5%), and Act (20%) gene. laf-A was missing in A. veronii and A. caviae isolates. Act and Hyl were absent in A. sobria isolates. The Aeromonas spp. isolates recovered from water also harboured Aer (17.5%), Ser (12.5%), laf-A (12.5%), ela (5%), Hyl (2.5%), and gcaT (2.5%) genes, while Act gene was missing in these isolates.
The aadA resistance gene was the most detected among Aeromonas spp. isolates obtained from fish (37.5%), followed by tetC (32.5%%), tetA (27.5%), sul 1 (20%), and blaTEM (10%). Aeromonas spp. isolates recovered from water harbored aadA (20%), tetC (15%), tetA (7.5%), and sul 1 (10%) genes, while blaTEM was not detected in any of these isolates.
Antibiotic susceptibility testing
All Aeromonas spp. isolates (100%) recovered from both fish and water were resistant to ampicillin 10 μg, amoxicillin 30 μg, and gentamicin 10 μg. Isolates were highly susceptible to ciprofloxacin 5 µg (47.5%), florfenicol (40%) and trimethoprim 1.25 μg/sulfamethoxazole 23.75 μg (32.5%).
Pathogenicity of Aeromonas spp.
Tilapia injected with Aeromonas spp. exhibited skin darkening, and hemorrhages on the external body surfaces, while some fish showed erosions and fin rot (Fig. 1). Control fish showed normal clinical signs. The LD50 value determined for the four tested aeromonad isolates was as follows: A. veronii (5.5 × 105), A. hydrophila (4.21 × 106), A. sobria (5.6 × 106), and A. caviae (8.48 × 106). Congestion and enlargement of the liver, spleen, and kidneys were the most common post-mortem lesions. Bacterial strains were reisolated from dead fish, and no mortalities were recorded in the control fish.
Histopathological observation
Degenerative, necrotic changes and mononuclear inflammatory cell infiltrations were the most commonly seen histopathological alterations (Fig. 3)

Discussion
Bacteriological and molecular examinations confirmed the identity of motile aeromonads involved in Nile tilapia mortality. The phenotypic and molecular characteristics of the isolates were consistent with previous findings (Abu-Elala et al. 2015). Outbreaks of MAS in the investigated farm was due to inadequate management and lack of biosecurity practices that negatively impacted the physiological and immunological status of fish, rendering them more susceptible to infections (Elgendy, Moustafa, et al. 2015; Elgendy, Soliman, et al. 2015). Untreated poultry manure acts as a source of infections (Abu-Elala et al. 2015).
The motile aeromonads isolated from fish and water displayed numerous virulence factors, including the production of extracellular enzymes that enable aeromonads to adapt to unfavourable environmental conditions and facilitated their invasion into fish (Chuang et al. 1997). The virulence genes of the aeromonads have an important role in their pathogenesis. A relatively similar virulence gene profile to the one seen here was reported by El-Gohary, Zahran, and Abd El-Gawad (2020).
The recovered isolates were resistant to most of the antibiotics tested and harbored numerous resistance genes similar to those reported by Ndi and Barton (2011). This dilemma is exacerbated by the contamination of fish farms with agricultural discharges and the use of untreated poultry manure. Aeromonas spp. are intrinsically resistant to ampicillin, amoxicillin/clavulanic acid, and cefazoline antibiotics (CLSI 2010). Application of good biosecurity measures, use of medicinal plants as alternative to chemotherapeutics, prudent usage of antimicrobials and performing antibiotic sensitivity testing are critical issues to reduce antibiotic resistance in aquaculture (Elgendy, Awad, et al. 2021; Elgendy et al. 2022).
The challenge experiment confirmed the pathogenicity of Aeromonas spp. isolates. The extracellular components and virulence genes of Aeromonas spp. are among the primary determinants of their pathogenicity (El-Gohary, Zahran, and Abd El-Gawad 2020).
Conclusion
Stressed fish lose their physiological condition and become vulnerable to motile aeromonad infections. Motile aeromonads harbour numerous virulence and antibiotic-resistance genes. Appropriate management and biosecurity practices are essential in aquaculture for protecting the health of farmed tilapia.
Conflicts of interest/Competing interests
All authors declare that they have no conflict of interest.
Authors’ contributions
This study was conducted in cooperation between all authors.
Funding
No fund
Ethics approval
The study followed the guidelines of the Institutional Animal Care and Use Committee, National Research Centre, Egypt under the number (231192022).