The South American lungfish (Lepidosiren paradoxa) is found in swamps and slow-moving waters of the Amazon and Paraguay rivers, as well as in the lower basins of the Paraná River in South America. This species possesses lungs, allowing it to exchange gases with the air, making it the sole member of the Lepidosirenidae family (Froese and Pauly 2014). The natural habitat of these fish disappears during the dry season, during which they hide in the mud and create a chamber-like burrow about 30-50 cm below the surface, leaving some holes on the surface for air entry (Paxton and Eschmeyer 1998). The presence of lungs in these fish is of significant interest in the phylogeny of the respiratory system in vertebrates. During the transition from aquatic to terrestrial respiration, various alternative systems emerged, such as nasal mucosa, labyrinth, intestinal mucosa, skin, among others (Wang et al. 2021). In general, we know that air-breathing can occur through any moist and permeable membrane, with cutaneous respiration being well-known in amphibians, and it is possible that in some fish, the lining of the mouth and pharynx may function in this way (Bassi et al. 2005). Although some authors have described the histology of the lung of L. paradoxa in detail, both with light and electron microscopy, a parallel with the structures and cells of vital importance for the lung to perform proper gas exchange (hematosis) has not been established (Hughes 1973). In other articles, the same authors conducted a comparative study of the ultrastructure of the lung in frogs, lungfish, and rats (Hughes and Weibel 1978).
Five adult specimens of Lepidosiren paradoxa weighing 318.06 ± 77.12 with a length = 21.16 ± 2.55 cm were collected in the National Center for Aquaculture Development (CENADAC), Corrientes Argentina and were taken to the Laboratory of Immunology and Pathology of Aquatic Organisms (LIPOA) from the Federal University of Rio Grande, RG. The specimens were anaesthetized with 0.5% benzocaine (Sigma-Aldrich) for approximately 30 minutes according to de Moraes et al. (2005). Later, a necropsy was performed following the protocol of our laboratory (Romano, Sardella, and Russomando 1987). Fragments of lung tissue were fixed in 10% buffered formalin, processed using an automatic processor (Leica TP1020), embedded in paraffin, and cut using a microtome (Leica RM2245). The samples were then stained with hematoxylin and eosin.
For electron microscopy, we followed a previously established protocol (Luchini, Wicki, and Romano 2015). Small 1 mm fragments of lung tissue were cut and immediately fixed in phosphate-buffered glutaraldehyde at 3 % (pH 6.9 at 4°C), washed in Millonig solution, and post-fixed in 1% osmium tetroxide. The tissue blocks were then dehydrated in a graded series of ethanol-acetone, immersed in propylene oxide, and embedded in Durcupan ACNI (Fluka Chemie A.G., Switzerland). Ultra-thin sections were cut using an LKB ultramicrotome and double-stained with uranyl acetate and lead citrate before being examined under a Jeol JEM-8T electron microscope (Jeol, 32, Tokyo, Japan).
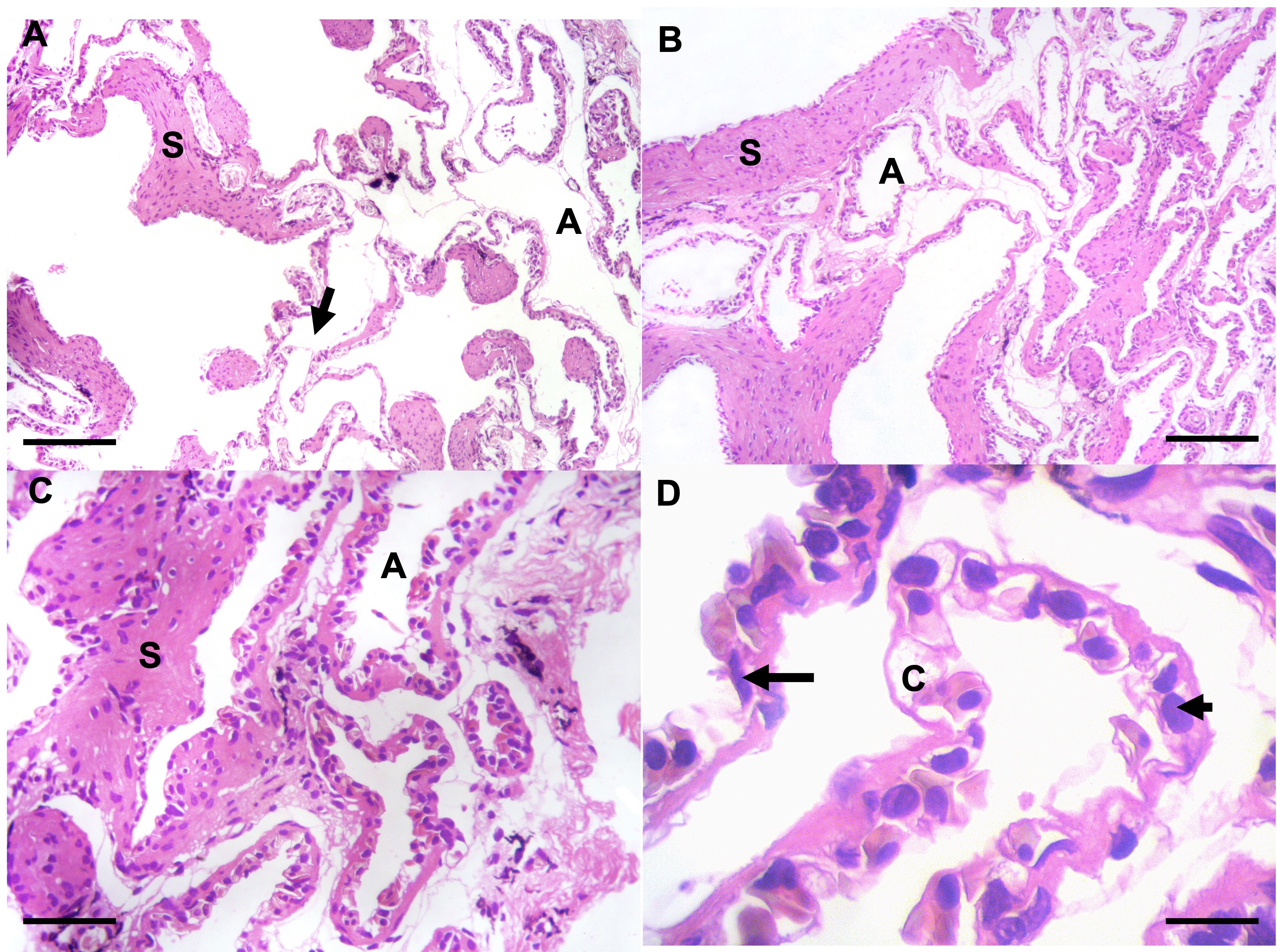
Macroscopically, no significant alterations were found. Macroscopy observation was performed to rule out any pathology that manifests macroscopically such as condensation, hemorrhage, among others. The objective is to ensure that the specimen studied does not present any pathology. In optical microscopy, the pharynx is lined with a non-ciliated mucosecretory pseudostratified epithelium (not shown). In the pulmonary structure, internal compartmentalization was observed. The lung is divided into compartments formed by septa or walls resulting from the folding of the walls with increasingly closed structures that form the alveoli. In some areas, structures resembling alveolar sacs that terminate in alveoli were observed, forming a spongy alveolar region. In the thin septa forming the alveoli, cells resembling type I pneumocytes with elongated nuclei lining the alveolar septa were observed. Additionally, larger cells with rounded nuclei resembling type II pneumocytes were also observed in the septa (Figure 1).
_with_alveoli_(a)_and_alveolar_d.jpeg)
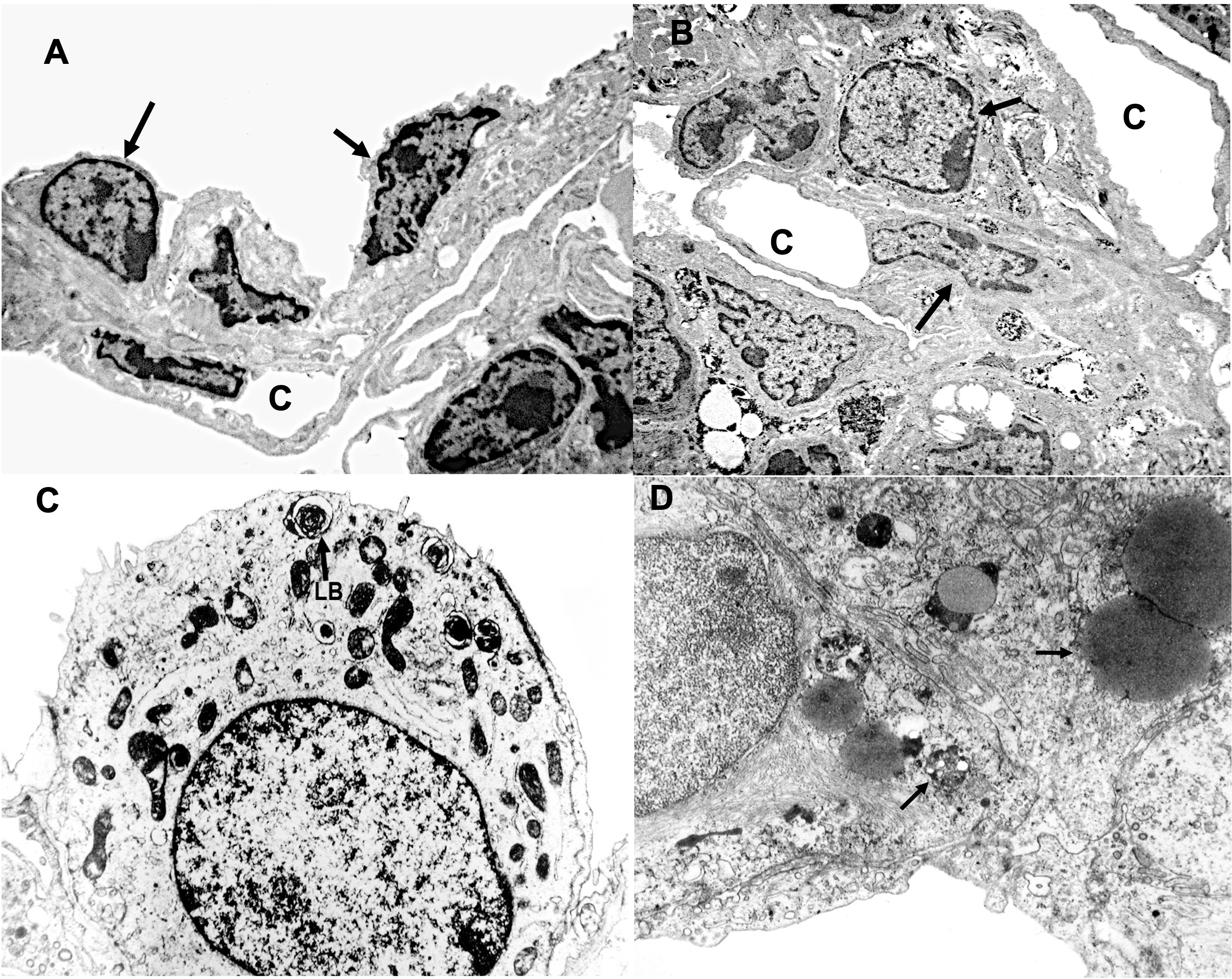
With electron microscopy, the details of the alveolar septum with different cells can be observed. Cells resembling type I pneumocytes with elongated nuclei and scarce cytoplasm are presente. We did not find significant structures such as pinocytotic vacuoles in the scant cytoplasm. These cells are associated with capillaries connected by their basement membrane, in close proximity to the respiratory medium, allowing for gas exchange.
Some cells resemble type II pneumocytes, with a central nucleus and multiple lamellar and osmophilic bodies in their cytoplasm, and blood capillaries run in this region (Figure 2).

We have not studied lungs belonging to dry season animals because some authors report that there are no differences (Icardo et al. 2017).
The most significant evolutionary change in Dipnoic fish is their possession of lungs as respiratory organs.. The gills receive blood that has already passed partially through the lungs. The heart’s atrium is divided into two chambers by a septum, and the ventricle is partially divided. Thus, the heart somewhat resembles the fully divided heart of mammals. In fact, the heart of Dipnoic fish exhibits a higher degree of structural division than that of any amphibian (Foxon 1955). The blood leaving the lung returns to the left atrium, while the right atrium receives blood from the general circulation. The partial division of the ventricle tends to keep the two bloodstreams separate, so that oxygenated blood flows to the first two branchial arches, supplying the head with relatively oxygen-rich blood. Therefore, the lungfish represents the beginning of a complete separation between the circulation to the lungs and the circulation to the rest of the body (Johansen, Lenfant, and Hanson 1968).
The lung of Dipnoic fish, like that of other vertebrates, develops as a ventral outgrowth of the lower part of the pharynx. In the Australian lungfish Neocerathodus, the single (unpaired) lung sac lies dorsal to the gastrointestinal tract and is a highly vascularized and moderately trabeculated structure. African and South American lungfishes (Protopterus and Lepidosiren) have a considerably more advanced lung structure, with much more internal compartmentalization (Icardo et al. 2017).
Neocerathodus relies almost entirely on its gills for the exchange of O2 and CO2 when submerged underwater, even with access to air (Mueller, Joss, and Seymour 2011).
In Lepidosiren paradoxa, 96% of gas exchange occurs through its lungs. However, much less CO2 excretion, around 43%, takes place through the lungs. The respiratory quotient for pulmonary gas exchange is low, at 0.45, compared to gill gas exchange, which is 6.7% (Johansen and Lenfant 1967).
The embryological origin of the lung as a ventral growth of the intestine is reflected in the striking similarity between their tissues and the unusual configuration of the airway and blood vessels. Internally, the lung is divided into compartments formed by septa of smooth muscle tissue resulting from the folding of its walls. These compartments are further subdivided to form a spongy alveolar region. In this region of increased surface area, blood capillaries run close to the respiratory environment, close enough to allow gas exchange (Zaccone et al. 2015; Jorgensen and Joss 2016).
The inflation of the lung is achieved by a pump-like action, as it is constituted by the hyoid apparatus and the muscular walls of the oral cavity. The exhalation of air is carried out by the contraction of smooth muscle components of the lung, aided by its natural elasticity provided by elastin fibers present in both the connective tissue and smooth muscle. The structure of lung of L. paradoxa is composed of partitions or septa with alveolar structures associated with capillaries ( da Silva et al. 2017; Glass 2011).
Unlike other authors, we observed a complex structure of the alveolar septum similar to that of mammals. The presence of septal lining cells with elongated nuclei and abundant cytoplasm, similar to type I pneumocytes, suggests that these cells are responsible for gas exchange. We also observed cells similar to type II pneumocytes with large round nuclei and osmophilic and lamellar material; these cells with their organelles may be related to surfactant production. The phylogenetic origin of these lungs is not entirely clear. However, the lung structure appears to be the most complex and adapted to aerial respiration, making it an important step between aerial and aquatic respiration.
Although some authors have described the histology of the lung of L. paradoxa in detail, both with light and electron microscopy, parallels with structures and cells of vital importance for gas exchange, i.e., for the lung to perform proper gas exchange, have not been established. In this study, we described the pulmonary morphology, which has been previously mentioned several times (Bernhard 2016), namely, the folding of septa to form alveoli. With electron microscopy, we described cells similar to mammalian type I pneumocytes (responsible for gas exchange) and cells similar to mammalian type II pneumocytes, with lamellar bodies and osmophilic bodies that may be associated with surfactant production. Surfactant forms a film at the interface between the pulmonary alveoli and the air, facilitating gas exchange.
This study was supported by research funds from MCT/CNPq - Project #301245/2016-09 MCT/CNPq/CT- Agronegocio/MPA Public Notice 036/2009 Project #308013/2009-3, Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), and Ministério da Pesca e Aquicultura (MPA).