
INTRODUCTION
Turbot, Scophthalmus maximus, is an economically important cultured flatfish species with fast growth, and high meat yield (Aydın et al. 2022). The main culture model for turbot is high-density aquaculture (Lei 2019). Flatfish production has reached 59,615 tons globally, mainly cultivated in Europe and China (FAO 2020). Turbot are increasingly being exposed to various stress factors such as extreme temperature fluctuations and deterioration of water quality due to the effects of global warming, and their welfare is under threat during their cultivation (Sun et al. 2022). Also, the dominance of high-density farming practices in turbot aquaculture, coupled with the lack of disease-resistant strains, has resulted in substantial economic losses due to severe outbreaks of bacterial, viral, and parasitic infections (Wang et al. 2010; Xiu et al. 2023).
The main bacterial pathogens causing furunculosis and edwardsiellosis in the turbot aquaculture industry are Aeromonas salmonicida and Edwardsiella tarda (Xiu et al. 2023). Other bacterial species such as Vibrio anguillarum, V. harveyi, and V. vulnificus are also known to cause infections in turbot (CSSA 2023). Before 2019, Edwardsiella piscicida, Aeromonas salmonicida, Mycobacterium marinum, and Streptococcus parauberis were identified as the predominant pathogens affecting turbot in China, where intensive aquaculture practices are prevalent (Gao et al. 2023). Furthermore, while rare, viral hemorrhagic septicemia virus (VHSV) has also been reported as a pathogen in turbot (Işıdan and Bolat 2011; Pereiro, Figueras, and Novoa 2016).
Lactococcosis is a significant threat to both cultured and wild fish populations worldwide. The disease manifests as hemorrhagic septicemia, causing high morbidity and mortality, and is resistant to antimicrobial treatment (Heckman et al. 2024). In recent years, lactococcosis has been recognized as one of the most prevalent diseases affecting farmed fish in both marine and freshwater aquaculture systems in Türkiye (Ture, Altinok, and Alp 2018). On the other hand, Lactococcus garvieae was initially recognized as the sole causative agent of lactococcosis. It has been reported to cause significant economic losses in various fish species causing hemorrhagic septicemia, particularly during the spring and summer when water temperatures exceed 16°C (Vendrell et al. 2006). Subsequently, a lactic acid bacterial strain isolated from a facial abscess in a sugar glider was genetically and phenotypically characterized. The strain was identified as a member of genus Lactococcus based on its morphological, physiological, and phylogenetic properties. Representing a new species within this genus, the bacterium was named Lactococcus petauri (Goodman et al. 2017). A L. garvieae strain, originally isolated from rainbow trout during lactococcosis outbreaks in Greece in 2007, has recently been reclassified as a new species, L. petauri (Kotzamanidis et al. 2020). Similarly, a whole-genome analysis of a Lactococcus garvieae isolate obtained from Türkiye years ago revealed that it was actually Lactococcus petauri (Altinok, Ozturk, and Ture 2022). In the following years, bacterial isolates from Türkiye, Spain, and Greece, predominantly identified as L. garvieae, were reclassified as L. petauri (Stoppani et al. 2023). Fish infected with L. garvieae and L. petauri exhibit similar clinical signs and cause significant economic losses in aquaculture due to high mortality rates, growth retardation, and treatment costs during outbreaks (Martinovic et al. 2021).
This study examined clinical signs such as loss of appetite, swimming disorder, and death in juvenile turbot fish. As a result of the research, L. petauri bacteria was reported for the first time in cultured turbot.
Material Methods
Background
All procedures involving turbot were conducted in compliance with National and Institutional Animal Welfare Guidelines. The research received ethical approval from the Local Ethical Committee of the Trabzon Central Fisheries Research Institute (SUMAE), Türkiye, under protocol number 325.04.02-19. The turbot examined in this study were cultured under SUMAE’s supervision in Trabzon, Türkiye, using a flow-through seawater system, with an annual production of approximately 5,000 adult fish. In late summer 2024, a health assessment was initiated in response to a subset of juvenile fish exhibiting anorexia, abdominal distension, and exophthalmos. Ten moribund or recently diseased turbot (weighing 50–60 gram) displaying clinical signs of disease were aseptically collected and immediately transported to the fish diseases laboratory for parasitological and bacteriological analyses. Sampling was conducted in late September 2024. The fish were stocked at an average density and fed with commercial feed. Water parameters were recorded as follows: temperature 18 ± 1°C, pH 7.4 ± 1, and salinity 18 ppt.
Parasitic examinations
The parasitological examination was conducted as described by Türe, Cebeci, and Özcelep (2022). Each fish was screened for the presence of ciliate species, which are prevalent in flatfish. Skin and gill scrapes were taken and analyzed microscopically under 20× and 40× magnifications using a Nikon E400 light microscope (Nikon, Japan).
Bacteriological investigations and biochemical identification
A macroscopic examination of the ten juvenile turbot focused on external areas such as the fins, ocular region, and skin. Internal organs, including the liver, spleen, and head kidney, were sampled and cultured on Tryptic Soy Agar with 1.5% NACI added (TSA; Merck, Germany) and Thiosulfate Citrate Bile Salts Sucrose (TCBS) agar (Merck, Germany). Cultures were incubated at 25°C for 48 hours. After incubation, bacterial colonies exhibiting smooth morphology were sub-cultured on TSA to ensure purity. Biochemical identification of pure bacterial isolates included Gram staining, cytochrome oxidase, catalase, and motility assays, following the methods of Ture, Altinok, and Alp (2018).
The sucrose fermentation test was also reported as an efficient differentiation method for L. garvieae and L. petauri strains (Saticioglu et al. 2023). L. garvieae (ATCC 49156) was used as a positive control. Bacteria were inoculated into a medium prepared by supplementing the OF basal medium (Merck) with 1% sucrose to detect carbohydrate fermentation. The yellow color that appeared after 48 hours in samples incubated under aerobic and anaerobic conditions was considered positive (Anonymous 2024).
DNA extraction, universal PCR assay, and sequencing
Genomic DNA was extracted from gram-positive bacterial isolates using the QIAmp DNA Mini Kit (Qiagen, Germany), by the manufacturer’s instructions. The concentration and quality of DNA were measured using a NanoDrop spectrophotometer (ND 8000; Thermo Fisher Scientific, USA), and the DNA concentrations were adjusted to 100 ng/μL.
The partial 16S rRNA gene of the bacterial isolate was amplified using the forward primer 63f (5′-CAG GCC TAA CAC ATG CAA GTC-3′) and reverse primer 1387r (5′-GGG CGG WGT GTA CAA GGC-3′) as described by Marchesi et al. (1998). PCR was carried out in a thermal cycler (Applied Biosystems, Germany) using AmpliTag Gold 360 Master Mix (Thermo Fisher Scientific, USA). PCR amplicons were visualized by electrophoresis on a 1.4% agarose gel in 1× Tris-Acetate-EDTA (TAE; AppliChem, USA) buffer at 100 V for 60 minutes and stained with ethidium bromide. DNA fragment sizes were determined using a 100-bp DNA ladder (Bioneer, Korea).
The PCR amplicons were sequenced on an ABI 3500 Genetic Analyser (Applied Biosystems, USA) with the BigDye Terminator Cycle Sequencing Kit v3.1). BioEdit (Hall 1999) was used for sequence alignment and editing. The sequences were aligned using the ClustalW algorithm (Thompson, Higgins, and Gibson 1994). Low-quality sequences were removed from the alignment. The generated sequences were uploaded to the GenBank database and compared with available sequences using the BLAST function. 16S rRNA gene sequence-based phylogenetic tree was constructed in Mega 11 (Tamura et al. 2011) using the maximum likelihood method with 1000 randomized bootstrap iterations. The most suitable nucleotide substitution model was chosen based on AIC (Akaike Information Criterion) and BIC (Bayesian Information Criterion).
Multiplex PCR assay
A multiplex PCR (mPCR) was performed to differentiate between L. garvieae and L. petauri using the multiplex PCR assay designed by Ustaoglu et al. (2024) which contains species-specific genes such as the gene encoding the DUF1430 domain-containing protein found exclusively in L. garvieae (204 bp), the Lichenan permease IIC component found exclusively in L. petauri (465 bp) and the teichoic acid biosynthesis protein F serves as an internal control for both L. garvieae and L. petauri (302 bp). Multiplex PCR was performed with Hot Start Multiplex PCR Master mix (NucleoGene, Türkiye). PCR mixtures containing L. petauri (Lg-Per) DNA and PCR reaction mixture without template DNA were used for positive and negative controls, respectively. The thermal cycling conditions were carried out as described by Ustaoğlu et al. (2024). Subsequently, the PCR products were visualized as explained above.
RESULTS
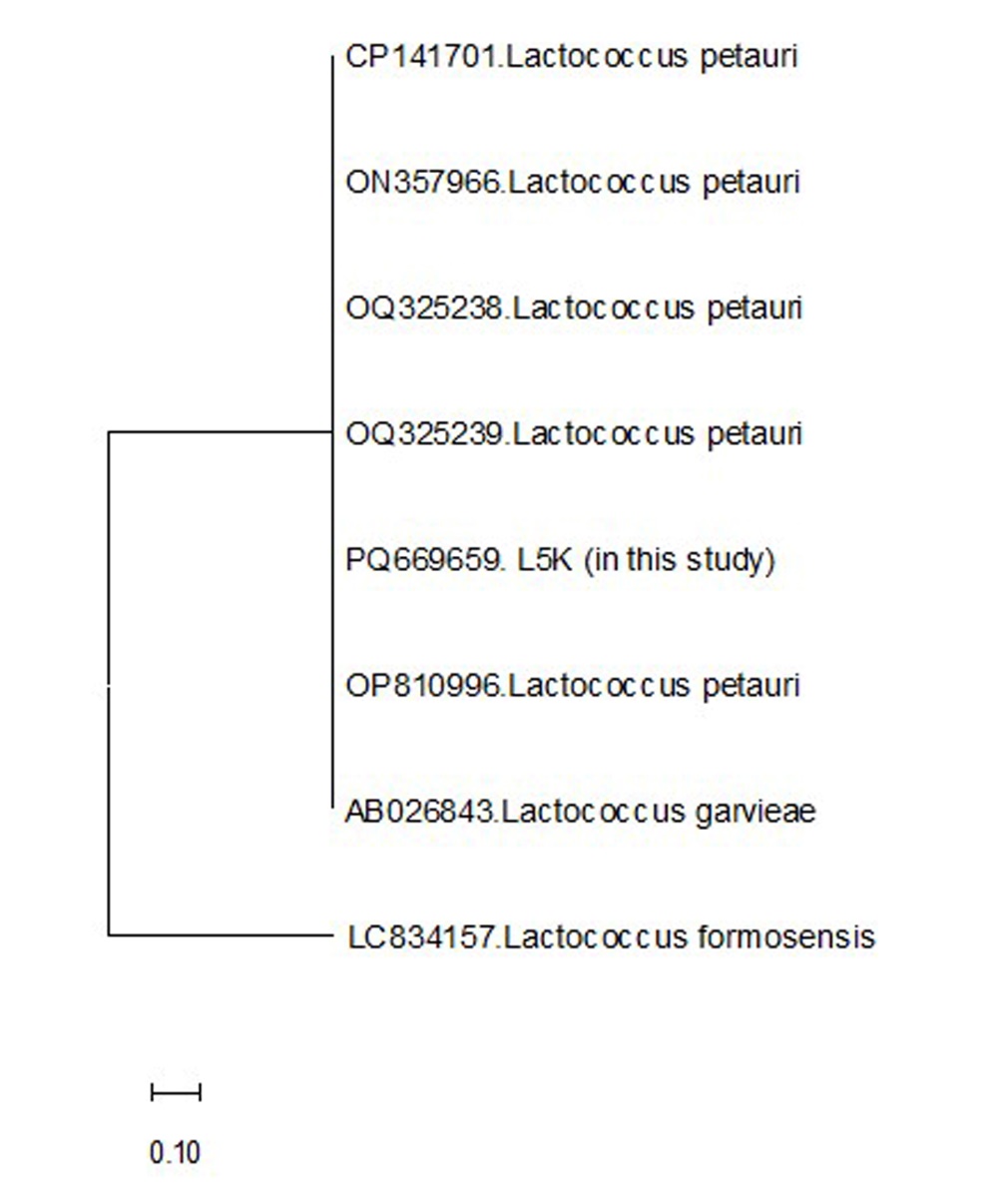
In the late summer of 2024, diseased turbot presented exophthalmia, redness around the ocular area, petechial hemorrhages at the base of fins, loss of appetite, and lethargy in a group of turbot. There was a substantial amount of yellowish fluid in the body cavity. Internally, the liver and spleen were also pale. During the study period, no parasitic agents were found in the turbot. Bacteria were isolated in all examined fish. Isolated bacteria were cocci or ovoid-cocci-shaped, gram-positive, and non-motile. Additionally, cytochrome oxidase and catalase showed a negative reaction. These biochemical properties resembled L. garvieae or L. petauri, which are frequently isolated from aquatic environments. The PCR amplification of the 16S rRNA gene from the bacteria yielded a product of approximately 1300 base pairs. Sequence analysis revealed that the isolate shares 99.56% similarity with the 16S rRNA gene sequence of L. petauri and L. garvieae in the GenBank database. The bacterium’s generated 16S rRNA gene sequence was deposited in GenBank under the accession number PQ669659. Moreover, the phylogenetic analysis based on the 16S rRNA gene sequence revealed that our isolate clustered closely with other L. petauri and L. garvieae strains (Figure 1).
_.jpeg)
DNA extracted from a presumptive Lactococcus spp. isolate and positive control (Lg-Per) was analyzed using a mPCR assay. Previously, NGS analysis revealed that L. garvieae Lg-Per strain was L. petauri (Altinok, Ozturk, and Ture 2022). The assay successfully amplified the targeted the Lichenan permease IIC component gene specific to L. petauri at 465 bp and the teichoic acid biosynthesis protein F gene, confirming L. petauri, at 302 bp. In contrast, DUF1430 domain-containing protein gene of L. garvieae at 204 bp was not detected (Figure 2).
__lane_1__*lactococcus_petauri*_pq669659__and_lane_2__*l.peta.jpeg)
Also, L. garvieae ATCC 49156 (control) isolate was negative for sucrose utilization, whereas the tested L. petauri isolate fermented sucrose in aerobic and anaerobic environments.
DISCUSSION
In recent years, lactococcosis has been identified as among the most common diseases affecting cultured fish produced in both marine and freshwater systems in Türkiye. A scientific study revealed that most of the L. garvieae isolates obtained from Türkiye were L. petauri (Altinok, Ozturk, and Ture 2022). Therefore, in this study, although the isolated bacterium was identified by sequence analysis, they were also confirmed by mPCR. This bacterium has been identified as the region’s most frequently isolated fish pathogen, underscoring its significant impact on aquaculture. To our knowledge, L. petauri was isolated from turbot for the first time. This study also highlights the importance of molecular tools in accurately identifying bacterial pathogens in aquaculture. Also, the confirmation of L. petauri as a dominant pathogen underscores its role in fish diseases in Türkiye.
The genus Lactococcus represents a heterogeneous group of lactic acid bacteria that play significant, yet sometimes contradictory, roles in human, animal, and food health. The fact that lactococcosis is caused by multiple species (L. garvieae, L. petauri, and L. formosensis) has been demonstrated by researchers worldwide through studies conducted over the past decade (Heckman et al. 2024). Lactococcus garvieae and the recently identified L. petauri are the causative agents of Lactococcosis, leading to significant losses in various cultured fish species, particularly at high water temperatures (Egger et al. 2023). Although 16S rRNA gene sequencing was previously regarded as the benchmark for microbial identification, its limited resolution makes it inadequate for distinguishing closely related species with high sequence similarity (Lan, Rosen, and Hershberg 2016). In contrast, whole genome sequencing offers greater precision in identifying and differentiating pathogens; however, it is expensive, time-intensive, and demands advanced expertise in bioinformatics (Cebeci et al. 2023). Previous studies have demonstrated that 16S rRNA-based PCR assays are insufficient for distinguishing L. petauri strains from L. garvieae strains, leading to frequent misidentification of L. petauri as L. garvieae (Altinok, Ozturk, and Ture 2022; Saticioglu et al. 2023). In the present study, BLAST analysis of sequence data from bacterial isolate, compared with reference sequences in the GenBank database, revealed 99%-100% sequence homology among the L. garvieae strains. A definitive diagnosis could be achieved using mPCR and a sucrose fermentation test.
In conclusion, this study represents the first report of L. petauri isolation from turbot. Additionally, mPCR and sucrose fermentation tests were effectively applied to differentiate L. petauri from L. garvieae strains.
Funding
This project was funded by the Republic of Türkiye, the Ministry of Agriculture and Forestry, General Directorate of Agricultural Research and Policies (Project no: TAGEM/HSGYAD/B/21/A5/P4/2505).
Conflict of interest
The authors declare that there are no conflicts of interest.
Author contribution
Esen Kulaç Polat and Mustafa Türe were responsible for the experimental design, analyses, and interpretation of the data. Atife T. Beken, Esen K. Polat, and İlyas Kutlu were responsible for the conduction of the experiment and for supplying resources. Mustafa Türe drafted the manuscript. All authors have read and agreed to the published version of the manuscript.
Data availability statement
The authors confirm that the data supporting the findings of this study are available within the article.
Ethical approval
Ethical approval was mentioned in the text.