Introduction
As VNN is an endemic disease in the Mediterranean, it can cause severe economic losses for aquaculture farmers and infect wild fish such as grouper, leading to high mortality rates and causing a decline in these endangered species (Bandín and Souto 2020). Viral nervous necrosis (VNN), also known as viral encephalopathy and retinopathy (VER), is an emerging neuropathogenic disease worldwide that affects a wide range of fish, both wild and farmed (Munday, Kwang, and Moody 2002; Savoca et al. 2021). It is caused by an RNA virus that belongs to the family Nodaviridae and the genus Betanodavirus (Mori et al. 1992; Comps, Pépin, and Bonami 1994), which affects nervous tissue, particularly the brain and retina, causing intense vacuolization of the granular layer of the brain, spinal cord, and retina, thereby leading to swimming abnormalities, lethargy, and darkening of the skin (Munday, Kwang, and Moody 2002; Doan et al. 2017; Zorriehzahra 2020; Fouzi and Sakajamary 2023).
Betanodavirus is a small, non-enveloped virus measuring approximately 25 to 30 nm in diameter and having an icosahedral symmetry structure. Its genome consists of a single-stranded positive-sense RNA divided into two segments, RNA1 and RNA2. RNA1 (3.1 kb) encodes the RNA-dependent RNA polymerase (RdRp), which is essential for viral replication, while RNA2 (1.4 kb) encodes the capsid protein, which is responsible for host tropism and immunoreactivity (Mori et al. 1992; Baud et al. 2015).
These viral particles are characterized by stability across a wide pH range (pH 2–9) and resistance to high temperatures of up to 56°C for 30 minutes, which enhances their persistence in the environment and their infectious potential (Frerichs et al. 2000; Bandín and Souto 2020; Kaplan et al. 2023). Betanodavirus has been classified into four main genotypes based on RNA2 variability: SJNNV (striped jack nervous necrosis virus), RGNNV (red spotted grouper nervous necrosis virus), BFNNV (barfin flounder nervous necrosis virus), and TPNNV (tiger puffer nervous necrosis virus), each adapted to specific thermal conditions (Nishizawa et al. 1995; Thiéry et al. 2012). The segmented nature of the genome facilitates reassortment between genotypes, commonly observed between RGNNV and SJNNV in Mediterranean fish populations (Toffolo et al. 2007; Baud et al. 2015; Toffan et al. 2016). Among these genotypes the RGNNV has the greatest host diversity in warmer water environments and the widest geographical distribution, covering Asia, USA, Australia, and the Mediterranean basin (Panzarin et al. 2012; Bandín and Souto 2020). On the other hand, RGNNV/SJNNV re-emergence has been reported in several Mediterranean countries (Biasini et al. 2022).
NNV disease was detected for the first time in the Mediterranean Sea (France) in farmed European sea bass Dicentrarchus labrax (Linnaeus, 1758) (Breuil et al. 1991) and then was reported for the first time in Algeria in 2011, during an episode of high mortality affecting wild adult dusky groupers Epinephelus marginatus and gold blotch grouper Epinephelus costae and later reoccurred in 2019 (Kara et al. 2014; Boukedjouta et al. 2020). These two species were long considered members of the Serranidae family, but now they are grouped within their own family, the Epinephelidae (Neubert et al. 2016; Tucker, Kurniasih, and Craig 2016). Epinephelus costae is a marine demersal fish found on sandy, muddy, or rocky bottoms, from the coastline to depths of 300 meters, it is found on the coasts of Italy, France, Spain, Tunisia, Algeria, as well as along the southern coast of Portugal and the eastern coast of Africa, from Morocco to southern Angola (Heemstra and Randall 1993). It is also found on the eastern and western coasts of Corsica. Epinephelus costae plays an ecological role in maintaining the balance of benthic ecosystems and is caught locally for human consumption or for recreational fishing. It is known for its organoleptic qualities and high commercial value (Bañón et al. 2018). Epinephelus marginatus is a nektro-benthic species with a discontinuous distribution divided into at least three main groups: the first in the northeast Atlantic Ocean and the Mediterranean Sea, and northern and northwestern Africa; the second in the southeaster Atlantic Ocean and southwestern Indian Ocean; and the third in the southwestern Atlantic Ocean (Schunter et al. 2011). Epinephelus marginatus is of high commercial value and it is highly prized by artisanal and recreational fishing (Smale 1986; Derbal and Kara 1996; Linde et al. 2004).
Epinephelus costae and Epinephelus marginatus are iconic species and are considered as symbols of marine protected areas (Bañón et al. 2018). Their populations are declining and they are considered endangered species on the Red List of Marine Fish of the Mediterranean Sea due to overfishing and their particular biological characteristics, including slow growth, long life span, large body size and protogynous hermaphroditism.
In this study, a virological and phylogenetic analysis was conducted to investigate and characterize the nervous necrosis virus in E. costae and E. marginatus, collected from different regions of the Algerian coast.
2. Materials and methods
2.1. Study area and sampling
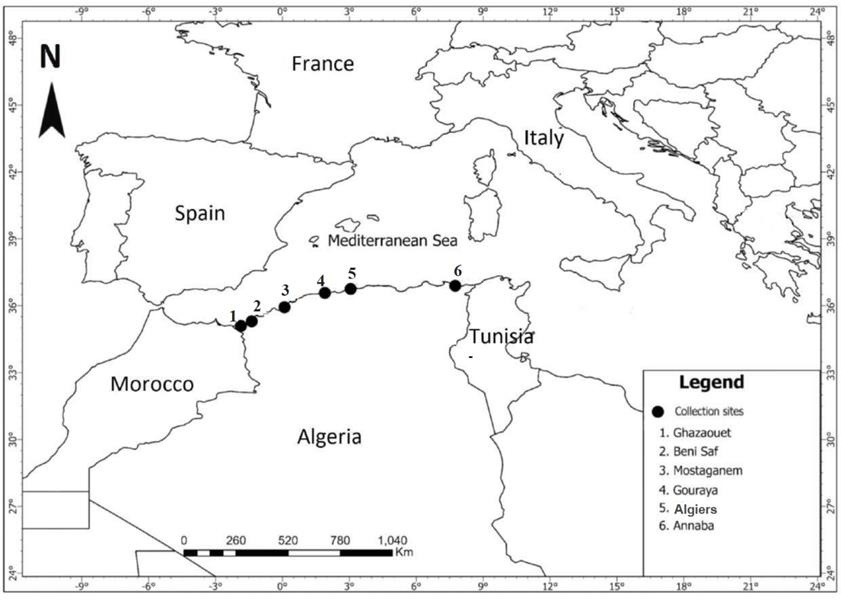
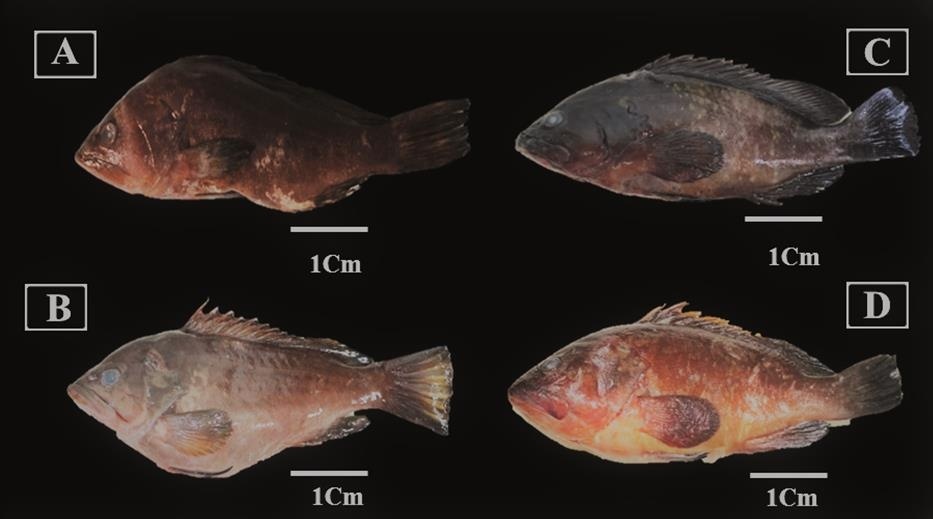
A total of 23 individuals of grouper were collected between May 2023 and July 2024 from various sites along the Algerian coast (Ghazaouet, Beni-Saf, Gouraya, Algiers, and Annaba) (Figure 1). All specimens were captured during fishing activity and noted to present cutaneous lesions and ocular opacity (Figure 2), with the majority collected during periods of elevated water temperature (around 25°C), a condition known to trigger betanodavirus activation and the onset of clinical signs associated with NNV disease (Toffan et al. 2016).
_of_*epinephelus_marginatus*_and_*epinephelus_costae*_alon.jpeg)
_observed_in_groupers_sampled_from.jpeg)
The heads of three fish (one specimen of Epinephelus marginatus, and two of Epinephelus costae) were collected and frozen at -20°C. The brains and eyes of twenty-one E. costae were collected and fixed in RNAlater. Part of the brain and one eye of E. costae were fixed in buffered formalin for histological and IHC investigations. Details of the biometric and geographic data of the samples are presented in Table 1.
2.2. Cell culture
The isolation of Betanodavirus was carried out according to standard protocol on the SSN-1 cell line derived from the head of the striped snake (Frerichs, Rodger, and Peric 1996; Toffan et al. 2019). The cell cultures were incubated at 25°C for 10 days and were monitored every 3 to 5 days to detect the appearance of the typical cytopathic effect. Three passages, each lasting 10 days, were performed before consider the sample negative.
2.3. Histology
The samples were dehydrated using a series of ethanol and xylene and embedded in paraffin. Sections of 3 µm were first deparaffinized, rehydrated, and then stained with Mayer’s hematoxylin-eosin (H&E) (Toffan et al. 2019).
2.4. RNA Extraction
Starting from frozen or RNAlater-fixed brain and eye samples, the homogenate was obtained manually using sterile quartz and the same medium used for cell culture (MEM 2414 - Sigma), then centrifuged for 15 minutes at 4,000 x g, at 4°C. RNA was extracted from 100 µl of homogenate (brain or retina) using the RNeasy Mini kit (Qiagen) according to the manufacturer’s instructions (Toffan and Panzarin 2020).
2.5. RT-qPCR
RT-qPCR reactions were performed in 25 µl with 600 nM of each primer, 400 nM of probe, 5 µl of RNA and 1× of Quantitect Probe RT-qPCR master mix (Qiagen) and the protocol was carried out using a CFX96 thermocycler (Biorad) with the following thermal profile: 30 min at 50°C followed by 15 min at 95°C and 40 cycles of denaturation at 94°C for 15 sec and 60 sec at 60°C for hybridization as described by Baud et al. 2015.
2.6. Sequencing, genotyping, and phylogenetic analysis
The genetic characterization of positive samples was obtained by reverse transcription followed by PCR amplification targeting the RNA-dependent RNA polymerase (RNA1) and the capsid protein (RNA2). The primer pairs used for the amplification of partial sequences of RNA1 and RNA2 were VNNV5/VNNV8, 521-For/VNNV6 and VNNV1/VNNV2 respectively (Toffolo et al. 2007; Bovo et al. 2011). The PCR were performed in 25 µl of total reaction volume with one-Step RT-PCR Kit (Qiagen), with a 400 nM concentration of each primer. The thermocycling conditions were as follows: 50°C for 30 min, 95°C for 15 min, followed by 40 cycles consisting of 40 sec of denaturation at 94°C, 40 sec of hybridization at 55°C, and 70 sec of elongation at 72°C; the reaction ended with a final elongation of 10 min at 72°C.
The PCR products were analyzed to verify their purity and size by electrophoresis on a 2% agarose gel, after staining with GelRed™ (Biotium), and then purified with the ExoSAP-IT® (USB) kit. The amplicons were sequenced in both directions using the same primers as those used for RT-qPCR, using the Big Dye Terminator v3.1 sequencing kit (Applied Biosystems). The products were purified using the PERFORMA® DTR Ultra 96-Well Plate kit (Edge BioSystems) and then sequenced on an ABI PRISM® 3130xl 16-capillary genetic analyzer (Applied Biosystems). The sequence data were assembled and edited using SeqScape® v2.5 software (Applied Biosystems). The partial sequences of RNA1 and RNA2 were aligned and compared to the corresponding regions of representative Betanodavirus strains available in GenBank which were selected based on sequence quality, completeness, and their relevance to the geographical and phylogenetic context of the present study, using the MEGA 7 software (Kumar et al. 2018). Phylogenetic trees based on partial nucleic alignments were also developed for both genetic segments using the neighbor-joining (NJ) method with 1000 bootstrap re-samplings available in MEGA 7.0 package.
3. Result
3.1. Cell Culture and Histology
The samples tested in cell culture (25ITT-V/70; 25ITT-V/71; 25ITT-V/72; 25ITT-V/86; 25ITT-V/92) did not show any cytopathic effect and were therefore considered negative. Furthermore, the histological sections of the tested samples (25ITT-V/75; 25ITT-V/77; 25ITT-V/81; 25ITT-V/85; 25ITT-V/86; 25ITT-V/88; 25ITT-V/89; 25ITT-V/93; 25ITT-V/94) did not reveal any vacuolar lesions or other typical signs (i.e. gliosis, congestion, etc.) affecting the retina and the brain (Table 2).
Some samples resulted positive at real-time PCR but negative at viral isolation on cell culture and histology because PCR is more sensitive and detects the presence of genetic material (DNA or RNA), whether it comes from a living, inactivated, or even partially degraded agent. Cell culture and histology can confirm viability and pathological effect. However, due to long samples storage these latter characteristics were lost in some samples.
3.2. Real-time PCR and sequencing
A total of 38 samples (brains and eyes) from the collected groupers were tested using RT-qPCR for the diagnosis of betanodavirus. Seven samples tested positive (25ITT-V/75; 25ITT-V/77; 25ITT-V/80; 25ITT-V/84, 25ITT-V/86; 25ITT-V/89 and 25ITT-V/92) as reported in Table 2. Four of them tested positive from brain only (25ITT-V/75; 25ITT-V/77) or from eyes only (25ITT-V/80; 25ITT-V/92), while three tested positive in both brain and eye (25ITT-V/84; 25ITT-V/86 and 25ITT-V/89). All samples were subjected to genetic characterization through the amplification of partial segments of RNA1 and RNA2. Unfortunately, and probably due to the long storage of the samples and consequent RNA degradation; only 4 samples’ sequences were obtained out of 7 positive. Sequences were submitted to GenBank and associated with the accession numbers from PV958793 to PV958800.
3.3. Phylogenetic analysis
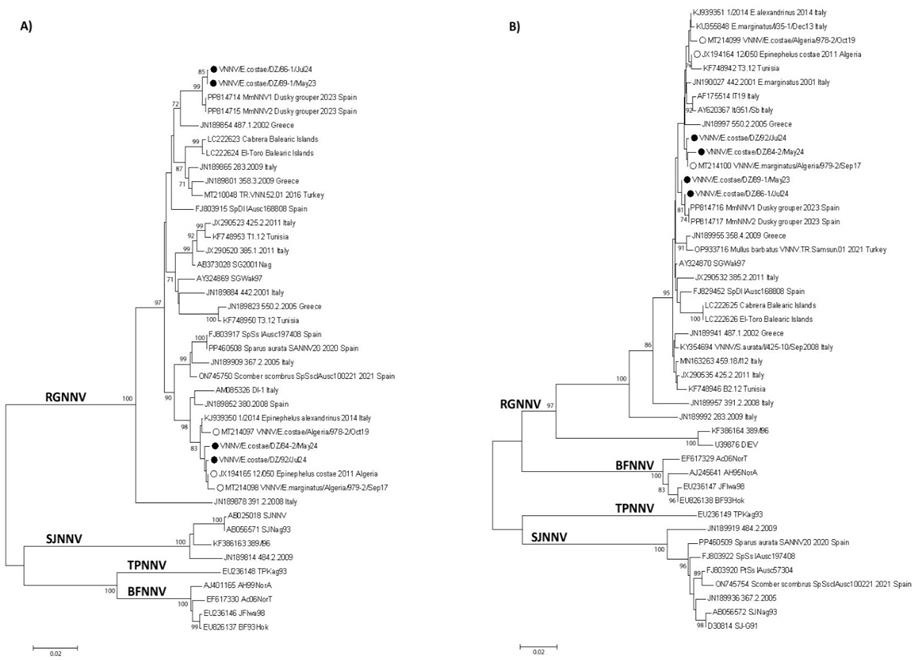
The Neighbour-joining (NJ) phylogenetic tree deduced for the partial sequences of RNA1 and RNA2 segments revealed that they all belong to the RGNNV genotype and the sequences obtained in this study are located within the coding regions and have slightly different lengths, depending on the strain, they were grouped with betanodaviruses from different Mediterranean countries previously reported in others studies (Figure 3).
According to the nucleotide sequences of RNA1 which corresponding to amino acid position from 43 to 352 of the RNA-dependent RNA polymerase with an average length of 930 nucleotides, the newly isolated viruses were grouped in two separate branches of the three; the first one closely related with previously reported for the Algerian Betanodavirus [2011 (Kara et al. 2014) and 2017 (Boukedjouta et al. 2020)] and the second one closely related to more recently detected Spanish Betanodavirus [2023 (Kersting et al. 2024)].
According to the nucleotide sequences of RNA2 which corresponding to amino acid positions are from 118 to 295 of the capsid protein with an average length of 534 nucleotides, similar correlation as for RNA1 was observed although with less strong statistical support (Figure 3).
_phylogenetic_tree_based_on_nucleotide_sequences_of_the_rna1_(a)_and.jpeg)
4. Discussion
The nervous necrosis virus is endemic to the Mediterranean basin, and it has been the subject of many scientific reports, both in wild and in farmed fish population (Gomez et al. 2006; Kara et al. 2014; Moreno et al. 2014; Vendramin et al. 2016; Valencia et al. 2019; Bandín and Souto 2020). Since the first mortality cases reported in 1999 in Ustica, Italy (Marino and Azzurro 2001), Betanodavirus infections in groupers have increasingly been documented across the Mediterranean.
In the present study RT-qPCR, and genome sequencing were used to screen and identify Betanodavirus genotypes found in fish collected from various sites along the Algerian coast. We used published primer pairs targeting both RNA1 and RNA2, which enabled us to identify potential homologous and reassorted betanodaviruses present in Mediterranean waters to date (Dalla Valle et al. 2001; Thiéry et al. 2004; Cutrín et al. 2007; Panzarin et al. 2010; Bandín and Souto 2020). Phylogenetic analysis using the neighbor-joining method of the two segments RNA1 and RNA2 revealed that both segments belong to the RGNNV genotype. This genotype is the most widespread in the Mediterranean region (Munday, Kwang, and Moody 2002; Panzarin et al. 2012; Costa and Thompson 2016; Toffan et al. 2017; Berzak et al. 2019; Bandín and Souto 2020; Kaplan et al. 2023).
In the Mediterranean, the reproductive period of groupers occurs between June and September. NNV was detected in two E. costae individuals from central Algeria (Algiers) and two others from western Algeria (Mostaganem), all collected during the post-reproductive phase. This period is characterized by physiological weakening of the fish, making them more susceptible to pathogens and environmental stressors (Kara et al. 2014). The finding of positive fish in two consecutive years (2023 and 2024), infected with viruses closely related and similar to those already detected in the same area in previous surveillance activity confirms once more the continuous viral circulation in the Algerian Epinephelus population.
On the other hand, the detection in 2024 of two strains clustering separately from the Algerian viruses, namely from Mostaganem, and closely related to Spanish recent isolates, suggest the introduction of a new strain of Betanodavirus from abroad. The spread of Betanodavirus in the natural environment is probably the result of marine commerce (trade exchanges already exist between the two countries, particularly facilitated by the maritime route linking Valencia to Mostaganem, which enables the movement of passengers and potentially of goods) or from neighbouring countries through the introduction of farmed fish populations. In addition, wild migratory fish can carry the virus to various geographical areas (Munday, Kwang, and Moody 2002; Berzak et al. 2019; Zrnčić et al. 2022).
Invertebrates, such as bivalve molluscs (mussels, clams, and oysters), can also serve as natural reservoirs and potential carriers of the VNN virus and, therefore, play a role in viral transmission in the water column (Volpe et al. 2018). Additionally NNV is highly tolerant to various environmental conditions and can survive for long periods in seawater (Frerichs et al. 2000). The infection disease spreads easily among healthy fish through horizontal transmission, particularly when the groupers share an environment with infected fish or are exposed to a suspension containing the virus (Shetty et al. 2012). Similarly, NNV spreads from resistant species introduced to susceptible species such as groupers (Boukedjouta et al. 2020).
Groupers are extremely susceptible to NNV, and outbreaks of VNN disease has been reported in several part of the Mediterranean basin: Spain, Italy, Greece, Tunisia, Algeria (Vendramin et al. 2013; Kara et al. 2014; Cherif and Fatma 2017; Bitchava et al. 2019; Valencia et al. 2019; Boukedjouta et al. 2020; Kersting et al. 2024). The fish belonging to this genus are therefore threatened not only by fishing activity, but also by viral infection. These constants threats coupled with the ecological peculiarities of the species, such slow grow rate, late reproductive maturity and protogyny put the survival of these species at risk. For these reasons, it is necessary to increase the establishment of marine protected areas in other regions of the Mediterranean Sea, which would be very beneficial in helping to conserve these species (Boukedjouta et al. 2020). In addition to implementing appropriate surveillance strategies to monitor the impact of the nervous necrosis virus and other infectious diseases, it is necessary to monitor and limit possible inter-species viral exchange with farmed fish, particularly from imports.
Finally, an international network should be created to improve and harmonize all conservation activities in the field of grouper protection in the Mediterranean as well as improving the diagnostic capacity of different countries laboratories to allow an early detection and therefore a better management of new outbreaks (Vendramin et al. 2016; Muniesa et al. 2020; Volpe et al. 2023; Toffan et al. 2025).
Acknowledgments
K.M. would like to express his sincerest thanks to the laboratory staff of the Istituto Zooprofilattico Sperimentale delle Venezie for their help and collaboration during the brief research stay of S.B. for the accomplishment of the current study, part of the first author’s PhD thesis under the supervision of KM. S.B. would like to thank the PhD students from the PVCMRMS lab. (Ahlem Benarous, Hanane Zerroual and Meriem Abbad) for their help and support in preparing the samples. The first author wishes to express her deepest gratitude to her father (Mohammed Benmokrane) for his valuable assistance in collecting the grouper specimens. The authors would like to thank the two anonymous reviewers and the editor for their time and comments that helped to improve the manuscript.
Author Contributions
This study was conducted in cooperation between all authors; Sample collection and species identification, SB, KM; Histology and cell culture, AB, AT, SB; molecular and phylogenetic analysis, LB, PB, SB, KM; writing—original draft preparation, SB, KM, AT; writing—review and editing, SB, KM, AT, LB, PB, AB; Supervision, K.M and A.T; Interpretation of results, AT, LB, SB, AB, PB, KM.
Compliance and ethical standards
All authors declare that the findings of this study were based on fish sampled directly from landings at fishing ports. This article does not contain any studies with live animals performed by any of the authors.
Conflicts of interest
The authors declare that they have no conflicts of interest to disclose.
Funding
This work part of the first author’s doctorate (PhD) thesis was supported by the General Directorate for Scientific Research and Technological Development (DGRSDT) under the authorityof the Minister of high education and Scientific Research (MESRS—Algeria), and by Istituto Zooprofilattico Sperimentale delle Venezie (Padova, Italy).
Data availability statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.