1. Introduction
Turbot, Scophthalmus maximus (Syn. Psetta maxima), is an economically valuable demersal fish species belonging to the family Scophthalmidae which is widely distributed in the Baltic Sea, the Northeast Atlantic Ocean, the Mediterranean Sea, and the Black Sea (Firidin et al. 2020). The turbot aquaculture industry has rapidly increased in the last three decades in European countries and China (Aydın 2021) and effects of ploidy status of the turbot on growth performance is well studied (Aydın et al. 2022, 2021; Cal et al. 2006; Fernandez-Pato et al. 1996). Among other factors, infectious diseases represent a major threat to turbot aquaculture causing serious economic losses. Overstocking and stressful conditions of culturing practices are making fish more susceptible to diseases and turbot culture has not escaped this fate. In the last three decades, due to intensification of farming conditions, mortality and morbidity episodes caused by pathogens have become a severe problem for turbot industry worldwide (Pereiro, Figueras, and Novoa 2016; Lei et al. 2006; Nougayrede, Sochon, and Vuillaume 1990; Westerdahl et al. 1994; Brudeseth et al. 2005; Feng et al. 2020).
Aeromonas salmonicida, causative agent of furunculosis (Öztürk and Altinok 2014), is a pathogenic bacteria that poses a threat in turbot culture (Coscelli et al. 2014). A. salmonicida mainly affects both farmed and wild salmonids (Austin and Austin 2016) but it has also been isolated from other commercially important fish species including turbot (Lago, Nieto, and Farto 2012; Pedersen and Larsen 1996; Perez et al. 1996; Nougayrede, Sochon, and Vuillaume 1990). Experimental challenges of turbot with A. salmonicida have been performed previously to determine pathogenicity of the bacteria (Björnsdottir et al. 2005; P. Wang et al. 2020; Lago, Nieto, and Farto 2012).
Vibrio anguillarum (formerly Listonella anguillarum), a causative agent of vibriosis, is one of the main causes of mortality in cold and warm water fish species (Öztürk and Altinok 2014; Toranzo, Magariños, and Romalde 2005; Boran et al. 2013). V. anguillarum has been numerously reported as responsible for severe infections in turbot culture (Myhr et al. 1991; Larsen, Pedersen, and Dalsgaard 1994; Planas et al. 2005). Experimental challenges of turbot with V. anguillarum have been performed previously to determine pathogenicity of the bacteria (Planas et al. 2005; Chair et al. 1994; Grisez et al. 1996; X. A. Wang and Ma 2019).
Viral hemorrhagic septicemia virus (VHSV) is among one of the most threatening viruses causing mass mortalities in turbot especially during early life stages (Pereiro, Figueras, and Novoa 2016). The disease outbreaks have been reported from different farms worldwide (Skall, Olesen, and Mellergaard 2005; Schlotfeldt et al. 1991; Isidan and Bolat 2011). Turbot has been numerously subjected to experimental challenges with VHSV to assess the susceptibility of fish and pathogenicity of the virus (Brudeseth et al. 2005; Snow and Smail 1999).
Triploid fish are not considered as genetically modified organisms (Piferrer et al. 2000). Triploidy can be induced through different types of applications including chemical, pressure, and temperature shocks (Chalmers et al. 2017). Turbot have considerably lower survival rate in early life stages compared to other cultured fish species and cold-shock induction is the only known triplodization method with high survival rate (Piferrer et al. 2000). The desired outcome of triplodization is sterility which is followed by superior growth and provides disease resistance in most of the cultured species comparable to their diploid counterparts (Weber et al. 2013; Chalmers et al. 2017, 2016). In contrast, some studies highlighted that triploid fish being more susceptible to infectious diseases (Tildesly 2008; Hakoyama et al. 2001; Ozerov et al. 2010). Functional sterility through triplodization has been confirmed in turbot with superior growth after sexual maturity (Cal et al. 2006). However, little is known about immunity and disease resistance of triploid turbot. The resistance against commonly encountered disease agents in turbot aquaculture needs to be evaluated to assess the suitability of triploid turbot for aquaculture. The present study aimed to determine and compare the response of diploid and triploid turbot to experimental challenges with A. salmonicida, V. anguillarum and VHSV to provide a broader understanding of the virulence of these pathogens in triploid turbot hosts.
2. Materials and Methods
2.1. Fish
The experiments of the present study were approved by Erzurum Veterinary Control Institute (Approval number 24/2032). Fish used in the present study were offspring of turbot, Scophthalmus maximus, that originated from the Black Sea. The juvenile turbot used in the experiments originated from a single family. Experimental fish were produced according to Aydın et al. (2021). Shortly, eggs stripped from a single female were artificially fertilized with milt from two males and divided into two equal batches: one for the diploid group and other one for triploid induction. Half of the batch was subjected to triplodization. The cold-shocking technique was applied for triploidy induction (Piferrer et al. 2003).
Experiments were performed at the aquaculture unit of Central Fisheries Research Institute in Trabzon, Turkey. The initial average body weight and length of diploid and triploid groups were 24.60±8.91 g, 11.50±1.26 cm and 24.18±9.01 g, 11.48±1.16 cm, respectively. Triploidy status of turbot was assessed by measuring the large axis of erythrocytes under light microscope. The average long axis of erythrocytes of triploids and diploids were 14.9±0.3 µm and 11.5±0.16 µm, respectively. Fish were acclimatized to laboratory conditions prior to experiments. During the acclimation period fish were fed twice a day and held under a photoperiod of 12 h of light and 12 h of dark. Five fish from each ploidy group were used to detect non-specific immune response indicators prior to pathogen challenge tests. Whereas, a total of 1080 clinically healthy turbot (540 diploid and 540 triploid) were used in pathogen challenge tests.
2.2. Non-specific immune response indicators
2.2.1. Isolation of leucocytes
Five juvenile turbot (27.2±1.05 g, 11.4±2.22 cm) were randomly sampled for the experiment. The fish were killed with an overdose of tricaine methanesulfonate; MS222 (400 mg / L). The head kidney was aseptically removed and homogenised with a tissue homogeniser. Homogenised tissue was transferred to 15 mL tube containing 3 mL of Histopaq 1119 medium (Sigma) and 3 m of Histopaq 1077 medium (Sigma). The suspension was centrifuged at 7000 X G for 10 min at room temperature. The leucocyte layer was transferred to a new tube and leucocyte suspension diluted in L-15 medium (Leibovitz). Leucocyte suspension concentration was adjusted to 2x106 cells/ mL.
2.2.2. Phagocytosis assay
An aliquot of 300 µL of leucocyte suspension (2x106 cells / mL) was poured onto glass slides and incubated for 2 h at 15°C for attachment. Unattached cells were washed with L-15 medium. Fluorescent latex beads at a cell/bead ratio of 1:10 were poured on the slide and the slide was incubated for an additional 1 h. Suspension was washed with fresh L-15 medium and stained with Diff-Quick dye following the manufacturer’s instructions. Phagocyted cells were visualised and counted under a fluorescent microscope.
2.2.3. Serum lysozyme activity
Serum lysozyme activity was measured according to method described by Amar et al. (2000). Briefly, 25 µL of serum sample and 175 µL of Micrococcus lysodeikticus suspension (0.075%) in 0.1 M phosphate citrate buffer (pH 5.8) were added to plate wells in triplicate and mixed. The dilution of the hen egg white lysozyme (Sigma) standard in 0.1 M phosphate citrate buffer (pH 5.8) was prepared and place into wells. Turbidity changes in wells were measured at 450 nm using a microplate reader (µQuant, BioTek) for 5 min at 30 s intervals. The equivalent units of activity of each serum was compared against hen egg white lysozyme standard by using Deltasoft3-139F software (Biometallics, USA).
2.2.4. Superoxide anion (O2-) production in head kidney leucocytes
The nitroblue tetrazolium (NBT) reduction assay was performed to evaluate superoxide anion (O2-) production in head kidney leucocytes. For this purpose, 15 µL sample (2x107 cells / mL) was mixed with 15 µL of NBT (1 mg / mL) containing L-15 media and incubated for 1 h. Following the incubation, 400 µL of N,N-dimethylformamide was added into suspension. The suspension was centrifuged at 3000 X G for 10 min and the optical density of supernatant was measured at 540 nm.
2.2.5. Alternative Complement Activity
Alternative complement activity in diploid and triploid turbot serum samples was assayed using sheep red blood cells (SRBC). SRBC were washed twice in ethylene glycol tetraacetic acid-magnesium-gelatin veronal buffer (EGTA-MG2+-GVB) and the concentration was adjusted to 4x108 cells / mL in the same buffer. Aliquots of 100 µL of test serum was serial diluted with EGTA-MG2+-GVB. Serial dilutions ranged from 1:4 to 1:128. The test serum was dispensed into wells and 30 µL of SRBC suspension was added to each well and incubated at 18°C for 2 h. The mixture was centrifuged at 400 X G for 5 min at 4°C. Supernatant was transferred to a clean tube and absorbance was measured at 540 nm (Epoch Microplate Spectrophotometer, Biotek). The degree of hemolysis and the serum volume were plotted on a log–log axis, and the dilution of serum that gave 50% hemolysis was used for calculating the ACH50 (Analyses of complement activity) using the following formula:
where k is the amount of serum giving 50% hemolysis, r is the reciprocal of the serum dilution. Values of ACH50 were expressed as units per mL.
2.2.6. Total Immunoglobulin
Total serum protein in both diploid and triploid fish were determined with biuret method. Serum obtained from each fish was divided into two equal proportions. One of the sera was diluted with 100X %0.85 NaCl and used as a standard. The other serum sample (0.1 mL) was incubated for 2 h with the same amount of 12 % solution of polyethylene glycol (PEG) (Test sample). Following the incubations, samples were centrifuged at 5000 X G for 10 min at 4 °C. The supernatant was diluted with 50X 0.85% NaCl. Absorbance was measured at 540 nm. Total immunoglobulin was calculated using the following formula:
Total immunoglobulin (mg/L)=AtAs×Conc. of standard (mg/L)
Where At is the absorbance of test sample, As is the absorbance of standard
2.3. Challenge Test
2.3.1. Pathogens
A. salmonicida (ATCC: 51413), V. anguillarum (ATCC: 43305), and VHSV (ckc-3 isolate) were used in challenge tests. A. salmonicida and V. anguillarum were cultured in TSA agar at 25 °C for 24 h. A single colony of each bacterium was suspended in BHI broth and incubated overnight at 25 °C. Suspension was centrifuged at 5000 X G for 10 min to harvest bacteria. The pellet was resuspended in PBS and final bacteria concentration of 1.0 x 107 cfu / mL was adjusted by using a spectrophotometer at 540 nm. The BF-2 cell line was used to propagate the VHSV. TCID50 (tissue culture infective dose 50) titer of previously propagated virus stock was calculated in a 24 well plate of 80% confluent BF-2 cells at 16°C.
2.3.2. Challenge
The challenge tests was performed with 2x 3x 2x 3 (ploidy x pathogen x challenge method x replicate) factorial design for 45 days. Both diploid and triploid turbot were subjected to intraperitoneal (IP) and immersion challenges with a single dose of A. salmonicida, V. anguillarum, and VHSV separately in triplicate for 45 days. After an acclimation period, 1080 fish were randomly assigned to 36 x 200-L flow-through aerated tanks at a rate of 30 fish per tank. For the injection group, fish were infected by IP injection of 0.1 mL of pathogens (final concentration of 1x106 cfu / 0.1 mL for A. salmonicida and V. anguillarum, 102 DKID50 for VHSV). Negative control groups (3 x 30 fish / tank) were injected with 0.1 mL of PBS. For the immersion group, the water level was lowered to 20 L in each tank and fish were exposed to 1x107 cfu bacterial pathogen / mL water (105 DKID50 for VHSV). After one h of exposure, the water level was re-raised and water flow was restored.
Blood samples were collected from 5 fish in each group at 0, 7, 14, 21, and 30 days post-challenge. Blood serum was separated by centrifugation and stored at -20°C until analysis. During the experiments fish were fed once a day with commercial turbot feed. Dead fish were removed daily and subjected to routine bacteriological and viral examination to verify challenge specific mortality. Water temperature (18°C) and salinity (18 ‰) were kept constant throughout the experiment.
2.3.3. Preparation of antigen, antibody generation and ELISA
Blood sera were mixed with Freud’s adjuvant in equal proportion. Rabbits (n=3) were injected with 1 mL of the serum-adjuvant mixture intraperitoneally. Injection was repeated on 7th day post-injection with the same amount of adjuvant-serum mixture injection. Blood samples were collected from rabbits by cardiac puncture three weeks post-injection. Obtained hyperimmune sera were stored at -20°C.
From each sampling (0, 7, 14, 21 and, 30 days post challenge), 20 turbot from each of the infected groups and 5 turbot from the control groups were analysed. Plates were coated with 5 µg / mL of Poly-L-lysine diluted in 0.05 M carbonate buffer (pH: 9.6) and incubated overnight at 4 °C. A. salmonicida and V. anguillarum were cultured in BHI broth and incubated at 29 °C until bacterial optical density was reached to 0.4 (OD600). Bacteria washed with 100 µL PBS were added to each well. Plates were centrifuged at 5500 x G at 4°C for 30 min. Non-specific binding sites were blocked by adding 5% skimmed milk in PBS to all wells at 4°C overnight.
The wells were loaded with 100 µL of serum samples at two different dilutions (1:50 and 1:100) and incubated at room temperature for 2 h. Plates were washed three times with PBS containing 5% Tween (PBST). Wells were supplied with 100 µL of rabbit anti-turbot sera and incubated at room temperature for 2 h. Plates were washed with PBST, blocked with 2% goat serum, and incubated at room temperature for another 2 h, and rewashed with PBST. Wells were supplied with ethanolamine buffer (pH: 9.6) containing 1 mM MgCl2 and 100 µL of P-nitrophenyl phosphate substrate and incubated at room temperature for 45 min. Absorbance was measured at 405 nm in a plate reader (Epoch Microplate Spectrophotometer, BioTek).
2.4. Statistical Analysis
One-way analysis of variance (ANOVA) was performed to determine differences among challenge groups. Tukey’s HSD was used to test differences among group means for significance. Dunnett’s test was used to compare the groups against the control group. All statistical analysis were performed using R (ver. 4.0.4) with 95% confidence interval. Results are presented as mean±standard error of the mean.
3. Results
3.1. Non-specific immune response indicators
The phagocytic index and the percentage of phagocyted cells of diploid and triploid turbot were similar. Phagocytic index of diploid and triploid turbot were calculated as 2.32±0.87 and 2.47±0.96, respectively. The percentage of phagocyted cells of diploid and triploid turbot were calculated as 48.85±1.26% and 48.46±2.05%, respectively (Table 1). Superoxide anion production in the head kidney leukocytes of diploid and triploid turbot were calculated as 0.039±0.002 and 0.041±0.003, respectively. Serum lysozyme activity of diploid and triploid turbot were calculated as 686.2±17.33 u / µL and 456.00±9.56 u / µL, respectively. Diploid turbot had slightly greater lysozyme activity which is an indicator of non-specific immune response. Alternative complement activity of diploid and triploid turbot were calculated as 212.87±55.18 u / µL and 198.96±47.23 u / µL, respectively. Total plasma protein and total immunoglobulin values were calculated as 51.35±3.17 mg / mL and 13.17±0.82 mg / mL for diploids, 41.84±2.53 mg / mL and 13.17±0.82 for triploids, respectively (Table 1).
3.2. Challenge test
3.2.1. Aeromonas salmonicida
No mortality was recorded in diploid and triploid turbot following both immersion and intraperitoneal injection with A. salmonicida. Similarly, no mortality or abnormality were recorded in the control group.
3.2.2. Vibrio anguillarum
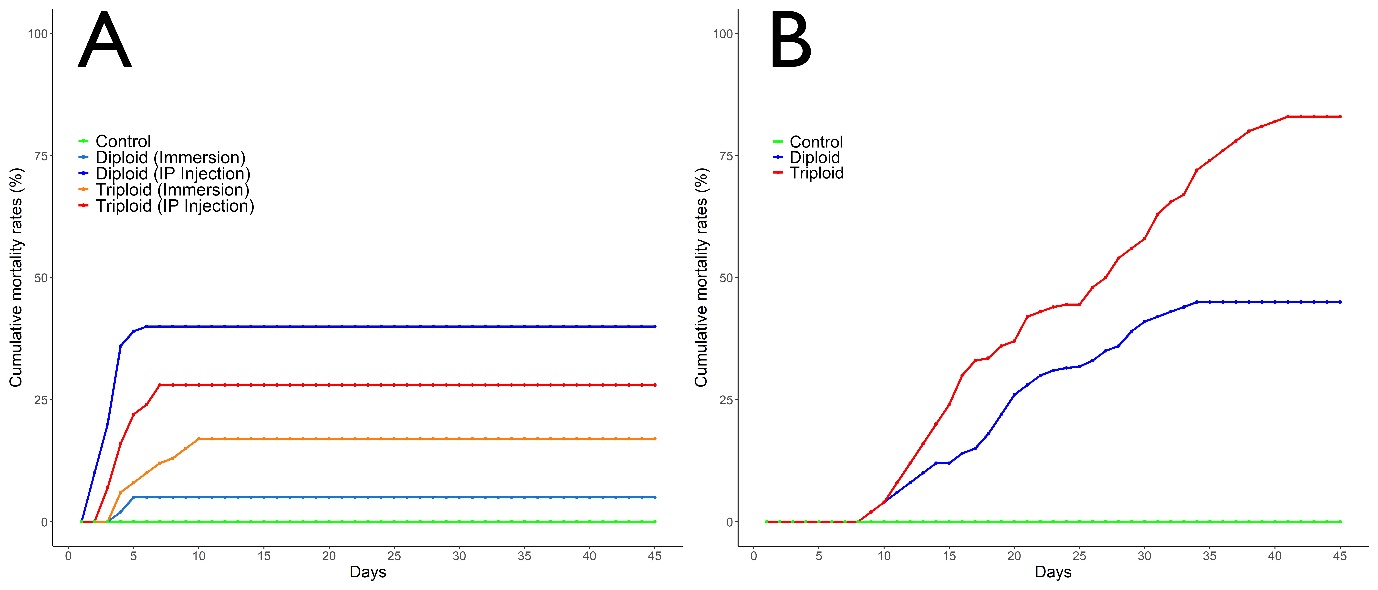
Fish begin to die on day 2 post-challenge in diploid and triploid groups following intraperitoneal (IP) injection with V. anguillarum and mortalities continued for the next 5 days. Cumulative mortality rates of diploid and triploid groups were calculated as 40.0% and 26.7%, respectively (Figure 1A). Mortalities began on day 3 post-challenge in diploid and triploid groups following immersion with Vibrio anguillarum and mortalities continued for the next 7 days. Cumulative mortality rates of diploid and triploid groups following immersion with V. anguillarum were calculated as 12.2% and 17.8%, respectively (Figure 1A). Challenge test results indicate significant difference in susceptibility against V. anguillarum between groups (P<0.05). The recovered bacteria from dead fish were verified as being V. anguillarum.

3.2.3. Viral hemorrhagic septicemia virus
Fish begin to die on day 9 post-challenge in the groups following IP infection with VHSV and mortalities continued until the day 36 post-challenge. Cumulative mortality rates of diploid and triploid groups were calculated as 53.3% and 86.7%, respectively. Typical symptoms of VHS disease was externally observed in moribund and dead fish subjected to IP injection. No mortality was recorded in ploidy groups following immersion with VHSV (Figure 1B). Challenge test results of triploid turbot showed an increased susceptibility against VHSV (p<0.05).
3.2.4. ELISA
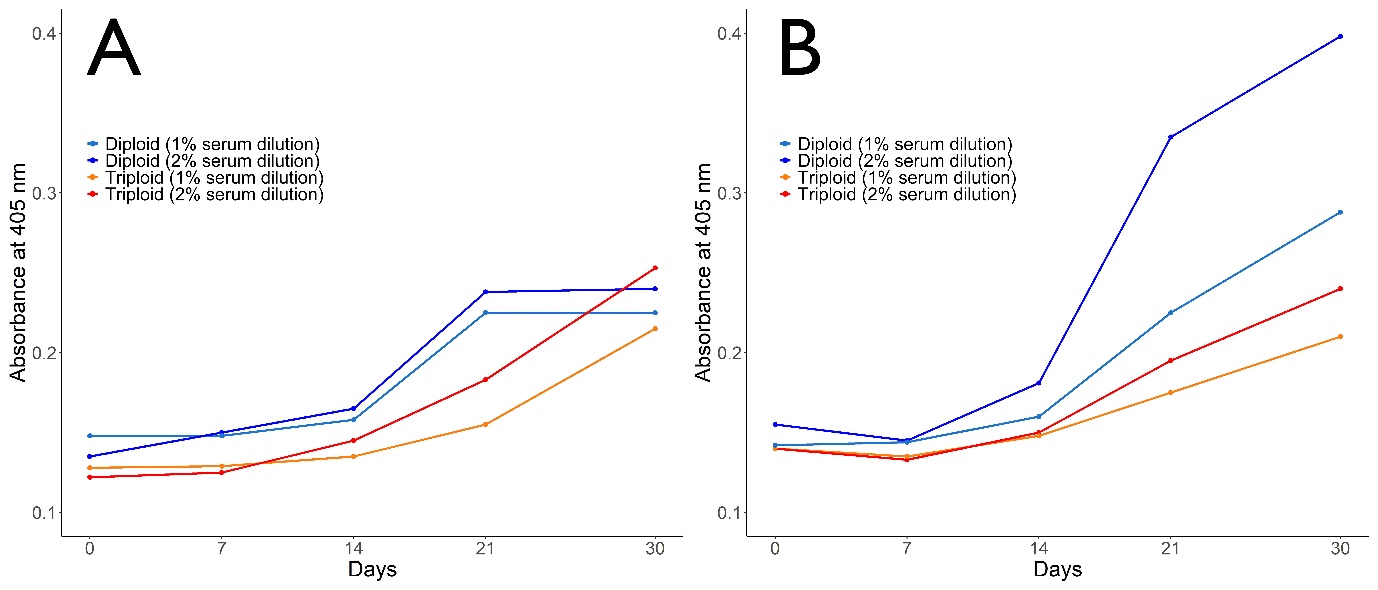
Specific antibody responses were observed at 1:50 and 1:100 serum dilutions (Figure 2). Antibody response against both V. anguillarum (Figure 2A) and A. salmonicida (Figure 2B) notably increased with each day after 14 days post-challenge at each dilution rates and peaked at 30 days post-challenge. There were no significant difference in terms of antibody response against V. anguillarum in both diploid and triploid groups. Antibody response of diploid turbot against A. salmonicida was significantly higher than triploid turbot (p<0.05). A notable increase in antibody response was noted after 14 days post-challenge until the end of the experiment in both diploid and triploid groups.
_against_*v._anguillarum*_(.jpeg)
4. Discussion
The main objective of this study was to compare innate immune response, disease susceptibility, and antibody response of diploid and triploid turbot following experimental infection with A. salmonicida, V. anguillarum, and VHSV. The present study is the first study that reports these infections in both diploid and triploid turbot using an immersion and injection route of infection.
Despite the clear advantages of being sterile (Aydın 2021; Piferrer et al. 2009) and superior growth after gonadal development (Cal et al. 2006), triploid turbot must perform similar or superior than diploids in terms of disease resistance and physiology in order to be considered for aquaculture. Triploids are commonly associated with the anatomic and skeletal malformities despite sharing almost identical external morphology (Maxime 2008). For instance, in triploid Asian catfish, Heteropneustes fossilis, the number of vertebrae were lower compared to diploids. In triploid Atlantic salmon, Salmo salar, high prevalence of skeletal deformity and reduced gill surface area was recorded (Sadler, Pankhurst, and King 2001). On the contrary, in demersal fish species, Hernanzez-Urcera et al. (2012) found a similar rate of skeletal abnormalities in diploid and cold-shock induced triploid turbot. Previous leukocyte profiles in diploid and triploid fish suggests that similar reactions would be developed during bacterial infection or stress (Maxime 2008; Svobodová et al. 2001; Kusuda et al. 1991). In the present study, some of the non-specific immune response indicators such as serum lysozyme activity and total immunoglobulin of triploids, were found to be comparatively lower than diploids which suggest that differences in response against pathogens might be developed based on ploidy status of turbot.
Turbot aquaculture is a growing industry and triploid turbot are now being used more extensively. Thus, elucidating how triploid fish would cope with economically important pathogens is important in determining their robustness and suitability to aquaculture. Previous studies indicate that culture performance of triploid fish is similar and sometimes superior than diploids (Fraser et al. 2013; Leclercq et al. 2011; Oppedal, Taranger, and Hansen 2003). But disease susceptibility varies depending on the pathogen strain, environmental factors, and origin of the species. Chalmers et al. (2017) challenged juvenile diploid and triploid Atlantic salmon, Salmo salar with Neoparamoeba perurans, the causative agent of amoebic gill disease, and ploidy appeared to have no significant effect on mortality rates. On the contrary, Powell et al. (2008) revealed that survival rate of triploid S. salar infected with N. peruarans was lower compared to diploids. Contrasting results may arise from different environmental conditions, challenge severity and origin of fish. Frenzl et al. (2014) revealed that the susceptibility of diploid and triploid S. salar against sea louse infection, caused by Lepeophtheirus salmonis, was similar.
A. salmonicida, causative agent of furunculosis, is known to cause high mortalities during disease outbreak in turbot aquaculture (P. Wang et al. 2020). Yet, no mortality was observed in both diploid and triploid fish subjected to challenge with A. salmonicida with IP injection and immersion. The A. salmonicida strain used in the present study was in vitro passaged a couple of times before challenge trials which might have resulted in alteration of virulence. Numerous previous studies have shown that the virulence levels of some of the bacteria are reduced during serial passage (Duangurai et al. 2020). Fresh clinical isolates might give different results. Our mortality finding contrast with an earlier study in which more than 50% cumulative mortality was observed in diploid turbot IP injected with 6.8 X 105 cfu A. salmonicida. The LD50 of different strains of A. salmonicida was estimated in juvenile turbot and ranged from 103 to 106 cfu / fish (Björnsdottir et al. 2005; Toranzo and Barja 1992). A final dose of 106 cfu / fish was IP administered to both diploid and triploid turbot, yet no mortality was recorded during the experimental period. The result of the present study indicates the A. salmonicida strain used in the present study as being a non-virulent strain.
In the present study, significantly higher mortality rates were detected following IP administration of V. anguillarum in both diploid and triploid turbot compared to the immersion method. It is an expected result since the IP administration is more invasive, bypassing natural barriers of infection and inoculating fish with high pathogen load. The cumulative mortalities of IP challenged triploid turbot were significantly higher than diploid counterparts which clearly highlights a difference in susceptibility between diploid and triploid turbot to V. anguillarum infection. Mortality rates of the IP infection with 109 cfu / mL V. anguillarum ranged between 23.33% - 75% depending on the fish family (X. A. Wang and Ma 2019). Juvenile turbot challenged with 106 cfu / mL of V. anguillarum caused 70% mortality within first 11 days (Planas et al. 2005). Hjelm et al. (2004) determined up to 80% mortality in juvenile turbot exposed to V. anguillarum via immersion at a concentrations between 103 and 107 cfu / mL. Although the injection with pathogen ensures delivery of a standardised dose, infection via immersion routes better reflect natural infections and outbreaks. Similar with the IP injection trial results, triploid turbot were more susceptible to V. anguillarum than diploids by immersion. The mortalities experienced during experimental infections with V. anguillarum are highly variable and unpredictable although the average level of mortality seen in the present study was comparatively lower than the reported range of mortalities. Water temperature, pathogenicity of the bacterial strain, genetic variation, and origin of the species might be the reason behind the differences observed in mortality rates followed by infection.
The VHSV causes disease in most farmed fish species including turbot and result in economic losses (Ross et al. 1994; Isidan and Bolat 2011; Öztürk and Altinok 2014; Schlotfeldt et al. 1991). In rainbow trout, similar susceptibilities were recorded in diploids and triploids against viral pathogens including IPNV, IHNV, and VHSV (Weber et al. 2013). Empirical evidence suggests that turbot infected with VHSV via immersion exhibit mortality rates around 70% (Snow and Smail 1999). In the present study, triploid turbot were found to be significantly more susceptible to the VHSV compared to diploids. The high level of mortality following infection with VHSV was comparable to the previous findings (Castric and de Kinkelin 1984; Snow and Smail 1999). The mortalities experienced during experimental infections with VHSV were highly variable, although the average level of mortality seen in the present study was within the reported range of mortalities.
5. Conclusion
Diseases are considered as the largest single cause of mortalities and economic losses in aquaculture and constrain the development of the industry. There are very few studies that have focused on triploid turbot health and immunity. Understanding how triploid turbot cope with pathogens is therefore important to evaluate their suitability to aquaculture. This study demonstrated that triploid turbot are more susceptible to V. anguillarum and VHSV infection than diploid turbot, providing vital information for the turbot aquaculture industry. A. salmonicida infection did not cause any mortality in ploidy groups via both IP injection and immersion. Having no mortalities during A. salmonicida infection in the present study might not reflect the true nature of the pathogen and its pathogenicity. Innate immune response of the ploidies were found to differ. Differences seen in immune response and disease susceptibility may be due to a compensatory mechanism for the reduced number of cells in triploids.
Funding information
This study was funded by Scientific and Technological Research Council of Turkey (TUBITAK) (Grand number 108R014).
Acknowledgement
The authors thank the personnel of the aquaculture unit of Central Fisheries Research Institute for their contribution and technical assistance during the experiments.
Conflict of interest
The authors declare that there are no conflicts of interest regarding the publication of this article.
Availability of data
The data that support the findings of this study are available from the corresponding author upon reasonable request.