_juvenile.tif)
Phaeohyphomycosis is a fungal infection caused by dematiaceous or melanised fungi (Seyedmousavi, Guillot, and de Hoog 2013). Although considered ubiquitous residents of plant material, soil, and wood, melanised fungi are likely adapted to specific niches that facilitate variable opportunistic or true pathogenic potentials. Exposure is typically associated with inoculation by minor trauma or inhalation. In mammals, infections are commonly cutaneous, subcutaneous, upper respiratory or primary cerebral, but in cold-blooded vertebrates are often disseminated and accompanied by severe tissue necrosis (Revankar, Sutton, and Rinaldi 2004; Seyedmousavi, Guillot, and de Hoog 2013).
In fish, various melanised fungi within the family Herpotrichiellaceae have been reported (Seyedmousavi et al. 2018). Fungi of the genus Cladophialophora have been only occasionally reported (Soto et al. 2017) and are considered commensals of gastrointestinal tract (Ghorbani-Choboghlo et al. 2014). In contrast, the genera Exophiala and Veronaea contain a number of well-recognized fish pathogens. Several Exophiala species have been associated with phaeohyphomycosis in various fish species (Seyedmousavi et al. 2018), but have never been reported in sturgeons. Conversely, Veronaea botryosa has been reported as the cause of systemic phaeohyphomycosis in two sturgeon species in the USA, the Siberian sturgeon (Acipenser baerii) and white sturgeon (A. transmontanus) (Steckler et al. 2014).
A juvenile beluga sturgeon (Huso huso), one of 11 mortalities from a group of 46 experiencing chronic losses while being grown for restocking purposes in a recirculating culture system, is the subject of this note. Diagnostic investigation of the outbreak identified varied microbial agents, including multiple bacterial species and a putative herpesvirus, as well as inconsistent pathologic findings suggestive of nutritional myopathy and this single case of phaeohyphomycosis. The mortalities were ultimately attributed to environmental stress and nutritional imbalances that likely predisposed the fish to a compromised immune system and opportunistic infections (Ciulli et al. 2020). The influence of temperature in this case is unclear, as the fish were being grown at 19°C near the high end of their 10-20°C optimal range (Froese and Pauly 2021).
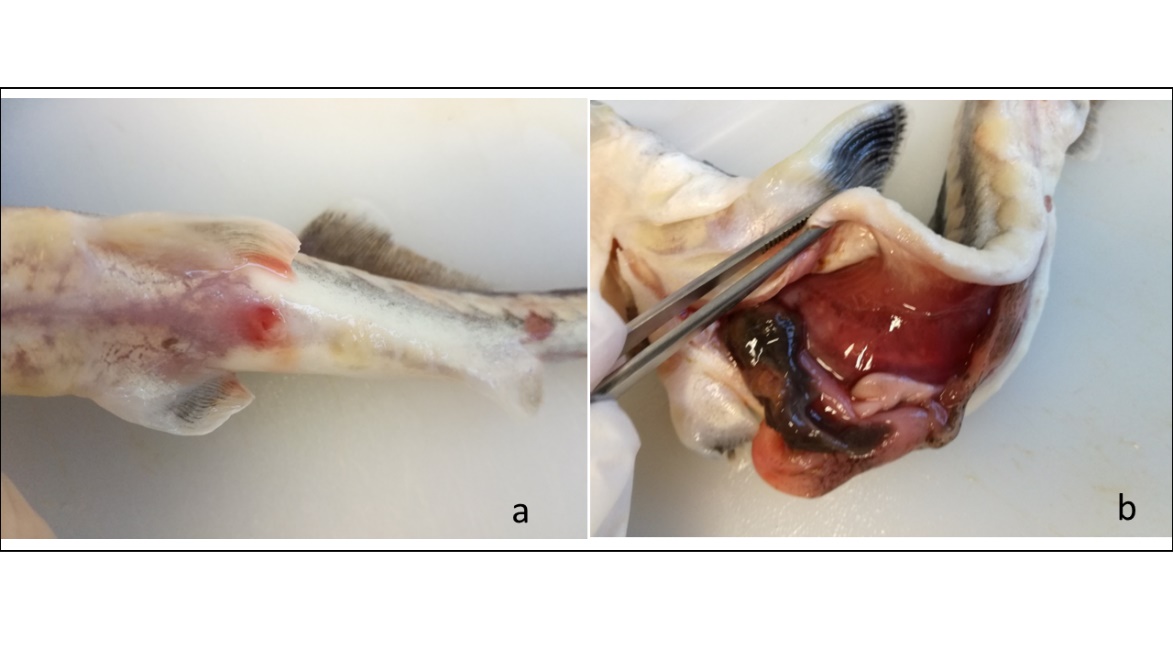
The fish was found dead in its tank and immediately transferred under refrigeration conditions to the diagnostic laboratory at the Local Operative Unit of Cesenatico (DIMEVET, Alma Mater Studiorum University of Bologna, Italy) for necropsy. Gross findings included arching of the body, multifocal reddening and erosion of ventral skin (Figure 1a), pale grey gills, sero-haemorrhagic coelomic effusion, a soft dark-brown liver, and a tan gastrointestinal tract devoid of ingesta (Figure 1b). Cytologic evaluation of coelomic fluid stained with May-Grünwald-Giemsa revealed pigmented, brown hyphae and bacteria later identified as Aeromonas veronii and Citrobacter freundii (Ciulli et al. 2020). In brief, a complete set of tissues (integument, head, gills, heart, liver, spleen, kidney, stomach, intestine) was collected, fixed in 10% neutral buffered formalin, processed routinely, and stained with haematoxylin & eosin (H&E) for histologic evaluation. Select sections were stained by the periodic acid-Schiff (PAS), Gomori methenamine silver (GMS), and Fontana-Masson (FM) methods. As the samples were collected some hours post-mortem, some autolytic findings, particularly at the level of intestinal mucosa, and some post mortem changes had occurred. However, after preservation tissues were deemed in a sufficiently good state for examination and interpretation. Pigmented, bronze hyphae, predominantly 3-5 µm (up to 10 µm) in diameter, with thin, non-parallel walls, infrequent septation, septal constrictions, and acute angle branching were widely distributed in all tissues examined. Hyphae were present transmurally in intestinal sections (Figure 2a) and were particularly abundant in the coelom adjacent to the intestinal serosa. Angioinvasion was common, with hyphae present in vascular lumens, including those of the gills and heart (Figure 2b). PAS and GMS stains highlighted the massive presence and morphologic features of hyphae in tissues (Figure 2c, d). The FM stain confirmed the presence of melanin pigment within the fungal walls (Figure 2e). Additional histological findings included severe lymphoid depletion of epicardial haematopoietic tissue, epaxial muscle pyrenophores degeneration and atrophy and multifocal ulcerative dermatitis.
_juvenile.jpeg)
_juvenile.tif)
In an attempt to identify the fungal agent, hyphae were excised from formalin fixed tissue sections using an MMI CellCut Plus laser capture microdissection (LCM) system (Molecular Machines & Industries, Glattbrugg, Switzerland) at the Department of Pathology, University of Georgia (Athens, GA, USA). Total DNA was extracted using a DNeasy Blood and Tissue Kit (Qiagen) after pre-treatment for formalin-fixed paraffin-embedded (FFPE) tissue following the manufacturer’s instructions. Whole genome amplification was then performed using a REPLI-g mini kit (Qiagen) to improve the quantity and quality of DNA. Endpoint PCR was attempted on both FFPE and REPLI-g amplified DNA using universal primers for amplification of fungal ribosomal RNA genes (White et al. 1990; Kurtzman and Robnett 1997) and a quantitative PCR (qPCR) performed following established protocols with V. botryosa specific primers (Yazdi et al. 2021) at the Department of Medicine and Epidemiology, University of California (Davis, CA, USA). However, corresponding products failed to amplify, likely the result of DNA degradation associated with formalin fixation. In a further attempt to identify the fungus, frozen samples including a DNA sample (20-25μL) and supernatant from a tissue lysate were sent to the University of California. However, due to shipping delays, the samples arrived at ambient temperature after 10 days in transit. The tissue lysate was centrifuged to produce a small pellet, the DNA extracted (Yazdi et al. 2021) then amplified and subjected to PCR and qPCR as described above. The presence of qPCR inhibitors was assessed by including an internal positive control for each sample. Again, no V. botryosa specific or other fungal products were detected. Control samples produced positive results as expected.
Despite the inability to identify the fungus by molecular methods, gross and microscopic findings in this juvenile beluga sturgeon are compatible with reports of V. botryosa-induced phaeohyphomycosis in other sturgeon species, including angioinvasion and widespread dissemination of pigmented hyphae (Steckler et al. 2014; Coleman et al. 2018; Groff et al. 2020). Significantly, the presence of coelomic fluid and involvement of serosal surfaces is typical of the condition known to producers of white sturgeon in the United States as “fluid belly” (Coleman et al. 2018).
Systemic mycosis by V. botryosa was the primary diagnostic consideration in this investigation due to association of the fungus with cultured white and Siberian sturgeon (Steckler et al. 2014; Coleman et al. 2018; Groff et al. 2020). However, additional novel and known pigmented fungi are increasingly identified as pathogens of bony and cartilaginous fish. Commonly implicated genera include Exophiala, Ochrochonis, and Phoma (Armwood et al. 2021). Significantly, fungal identification based on morphologic features, particularly in histologic sections, is unreliable and molecular methods should be employed when fresh or frozen tissue is available. Targets for identification based on PCR amplification and sequencing include the internal-transcribed spacer region of the rDNA (ITS), large ribosomal subunit gene D1/D2 domains (LSU), and the DNA polymerase II gene (PBP2) gene. While certain PCR protocols can work on FFPE tissues (Santos et al. 2013; Steckler et al. 2014; Meason-Smith et al. 2017; Cañete-Gibas and Wiederhold 2018), molecular characterisation attempts failed to identify the agent, presumably the result of DNA degradation due to formalin fixation and delayed sample processing caused by covid-19 associated shipping interruptions of frozen sample material.
Apart from the identification of the dematiaceous fungus, it is evident that the observed infection was predisposed by the poor health condition of the subject suggested by the bacterial coinfection findings, the immune cell depletion and the epaxial muscle pyrenophores degeneration. These detrimental effects are presumably due to nutritional imbalances and environmental stress induced by poor farming conditions. In these circumstances high pathogenic fungi such as Exophiala and Veronaea and less pathogenic ones such as Cladophialophora could take over the fish defence mechanisms.
Overfishing and poaching have greatly reduced wild populations of beluga sturgeon, prompting the IUCN to list the species as critically endangered (Froese and Pauly 2021). Declining numbers of wild sturgeon highlight the need for repopulation efforts based on artificial culture conditions. Despite the inability to identify the pathogen in this case, fish pathologists involved with sturgeon aquaculture should be aware of emerging fungal pathogens, particularly V. botryosa, as a potential threat to economic productivity and the conservation of beluga sturgeon, as well as culture conditions that may promote infection. Particularly, the setup of specific nutritional and environmental conditions are required for these ancient species even if this is actually limited by the paucity of information available (Hung 2017).
Concluding, fungal culture should be considered when sampling diseased sturgeon and fresh or frozen tissue is collected for use in molecular testing, to achieve an aetiological diagnosis and contribute significantly to the concurrent histopathological findings.