Introduction
Diseases induced by viruses count among the main threats and risk factors for aquaculture and for wild fish in fresh and brackish waters, but also in marine environments worldwide (Rodger 2016). Beside vaccination of fish (Biering et al. 2005), investigations were previously carried out to search for bacteria, algae or other agents from different environments with antiviral effects. Yoshida et al. (2013) found different bacteria with possible antiviral effects in gut contents of carp and koi. They also isolated ubiquitously occurring water bacteria identified as Pseudomonas ssp. which were able to inhibit at least koi herpesvirus (KHV). Other research groups investigated different components, e.g. exopolysaccharides (EPS) from bacteria (Gugliandolo et al. 2015; Biliavska et al. 2019) or marine algae (Reichert et al. 2017) for their antiviral properties. While the mode of action of EPS is still unknown or is only based on hypotheses, their efficacy has been demonstrated in different experiments. EPS from the cyanobacterium Arthrospira platensis, earlier known as blue algae, was obviously able to prevent the attachment of KHV to permanently cultured cells, especially to common carp brain (CCB) cells (Neukirch, Boettcher, and Sumrarn 1999). As a result, there was no or a very weak cytopathic effect (CPE) on the cells after infection with KHV when cells were treated with EPS (Reichert et al. 2017). Further on, Bergmann et al. (2020) reported that EPS can be used for prophylaxis of KHV disease (KHVD) or even as a metaphylactic measure during an outbreak to stop the disease.
Materials and methods
Algae (biomass (BM) and exopolysaccharides (EPS))
Algae (Arthrospira platensis NIES 39) had been centrifuged at low speed, dried, lyophilised and stored at 4°C until use. EPS were concentrated from the media and characterised as described by Reichert et al. (2016). The final EPS powder contained 0.4% pure polysaccharides. The EPS powder was stored at 4°C until use.
EPS in-vitro tests
EPS was additionally tested for its reaction with, or effect against, different fish viruses. Viruses used in the study were viral haemorrhagic septicemia virus (VHSV) from olive flounder (Paralichthys olivaceus) replicated onto Epithelioma papulosum cyprini (EPC, Fijan et al. 1983) cells, KHV replicated onto Common carp brain (CCB, Neukirch, Boettcher, and Sumrarn 1999) cells, reovirus obtained from marine environment (unknown marine crab) and viral nervous necrosis virus (VNNV) obtained from European sea bass (Dicentrachus labrax), both replicated onto E-11 cells (Iwamoto et al. 2000). All viruses with a titre between 10 and 100 TCID50/mL were mixed with the same volume of EPS solution between 16 and 100 mg/L in sterile phosphate buffered saline buffer (PBS) and stored at 4°C overnight. After incubation, the mixtures were 10 logs titrated using their original cell culture. After seven days, the titrations were evaluated. VHSV onto EPC cells was incubated at 15°C, all other viruses at 26°C.
Fish
For two experiments common carp (Cyprinus carpio, K1 age, n = 50 per group) were obtained from a commercial farm in the German federal state Thuringia. The farm (O. Fürneisen, Traßdorf) produces specific pathogen free carp. They are approved as free from spring viremia of carp virus (SVCV), KHV, carp poxvirus (CyHV-1) and carp edema virus (CEV). KHV and CEV were tested by quantitative real-time polymerase chain reaction (qPCR) (Bergmann et al. 2010; Bergmann, Klafack, et al. 2020), CyHV-1 by PCR and sequence analysis (Engelsma et al. 2013), and SVCV by reverse transcriptase-(RT)-PCR (Koutná et al. 2003). Additionally, 10 fish per group were investigated by KHV antibody enzyme linked immunosorbent assay (ELISA) to exclude an undetectable persistence of KHV (Bergmann et al. 2017). All fish were kept in tanks with a total volume of 480 L in a re-circulating system at 20°C with 12 h daylight and 12 h night. Daily 20% of the water (96 – 100 L) were exchanged, once a week 50% (240 L). After water exchange the required concentration of EPS was delivered daily to the aquaria to hold it in the water. For that reason, aliquots were prepared from EPS but also BM and stored at -20°C until their use. The fish were fed with commercial carp food four times a day.
Experiment 1 (BM and EPS)
The first experiment was performed with complete dried biomass (BM) of Arthrosphira platensis and purified, and dried EPS in order to examine if either of these two extracts can prevent the infection of the virus or at least the disease. The fish were challenged with KHV-E (European linage, Klafack et al. 2019). Four experimental groups were established in duplicates and one negative untreated control. Groups 1A and 1B were treated prophylactically for 14 days (d) with BM maintaining the EPS concentration in the water at 320 mg/L, groups 2A and 2B for the same period of time with 32 mg/L of extracted EPS. Groups 3A and 3B, 4A and 4B as well as the negative control group stayed untreated prior to challenge. During this time six fish from group 3A died because of, or due to, treatment against ectoparasites. At day 15 of the experiment, all carp were healthy and in good condition free from ectoparasites. At 16 d, the carp of groups 1A, 2A as well as 3A and 4A and the negative control carp, as the challenge control, were infected by immersion in a separate vessel with 104 TCID50/mL KHV-E in the water for 1 h. Subsequently, they were re-transferred back to their own tanks. After infection, the water of group 3A was additionally treated with BM, that of group 4A with EPS as a possible metaphylactic (therapeutic) measure, both in the same concentrations that were used for prophylaxis in groups 1A and 2A. The tank of group 1B was continuously treated with BM, that of 2B with EPS, both without infection. Groups 3B and 4B stayed untreated, serving as negative infection controls. All groups were observed for 30 days.
Experiment 2 (EPS)
In the second experiment the influence of prophylactic, metaphylactic, both or no treatment with EPS on the infection/disease development, were examined. In this set up 4 groups of fish were used: one control group without the addition of EPS (group 1), one group where fish were treated with 32 mg/mL EPS before and after infection with KHV (group 2), one group treated with 62 mg/mL EPS only prior to infection (group 3, prophylaxis group) and one group treated with 32 mg/mL only after infection (group 4, metaphylaxis). All groups were held for 6 weeks with/without EPS treatment prior to the infection with KHV. After 6 weeks, all groups were challenged using KHV-T (Asian lineage, Klafack et al. 2017) at a titre of 104 TCID50/mL in water by immersion for 1 h.
Serological investigation
At the end of both experiments, sera from 10 fish of each group were collected. All sera samples were examined by a validated KHV antibody ELISA (Bergmann et al. 2017) and by a newly established ELISA searching for antibodies against EPS. For the latter, EPS (10 µg/mL) diluted and adsorbed with PBS served as antigen on a Medisorp ELISA plate (Nunc, Germany). The adsorption was carried out overnight at 4°C. The next day, plates were blocked (1 h, room temperature, RT) using 300 µL Roti®Block (Roth, Germany) diluted 1:10 with distilled water. Next, plates were washed three times using PBS -Tween 20 (0.5%, PBST). Sera were serially diluted with PBST from 1/300 to 1/2,500 and incubated for 1 h (RT). After washing the plates three times again, a monoclonal antibody (mab) α-carp-IgM F 16 (Aquatic Diagnostics, Scotland) was used at a dilution of 1/64 with PBST as a secondary antibody. After incubation for 1 h RT, plates were washed three times. Further, the conjugate α-mouse-Ig-POD (Sigma, Germany) at a dilution (PBST) of 1/10,000 was added to wells (30 min, RT). Finally, the wells were washed again and 3,3’,5,5’-tetramethylbenzidine (TMB) substrate TMB (Sigma, Germany) was added to each plate. The reaction was stopped after 10-15 min at RT using 2 N sulfuric acid. The plates were read at 450 nm using an ELISA reader (BioRad, Germany). The cut off was adjusted and established at OD 0.2 using EPS negative sera (n=10) from carp.
Results
EPS in-vitro tests
Four different fish viruses were mixed and pre-incubated (24 h) with EPS solutions of concentrations ranging from 16 to 1000 mg/mL in order to test the potential for preventing the infection of the respective cells.
EPS were able to prevent cell entry of the investigated viruses at different concentrations (Table 1). While for 10 TCID50/mL KHV-T 16 mg EPS /mL prevented virus replication onto CCB cells, while for VHSV at 10 TCID50/mL 10 mg/mL EPS was already sufficient. Replication of the crab reovirus and the VNNV were successfully stopped when 32 mg/mL EPS was used. At higher concentrations than 128 mg/mL, EPS can be toxic to the cells and simulate a cytopathic effect onto cell cultures depending on their origin.
Experiment 1 (Biomass and EPS)
In this set up either dried BM of EPS were delivered to fish prior, and after the infection with KHV on day 14 or over the entire experimental period (30 days). The results showed that prophylactic delivery of BM and EPS to the tank water had no negative effects on the fish over 14 days. Before infection by KHV-E, neither clinical signs nor mortality induced by BM or EPS were found in the treated groups as well as in the two unchallenged negative control groups. However, it was observed that the oxygen concentration in the water dropped from 8 to 5 mg/L for 10 to 20 min after delivery of both substances into the water. In all KHV infected groups, KHVD was induced, though it ranged from weak to severe (Table 2). While in groups 1A and 3A, that were treated either prior to and after the infection, or only after the challenge with KHV respectively, the same mortality rates of 23 % were observed. In the first group (1A) the disease was unexplainably much more severe than in the second one (3A). For comparison, in the challenge control (negative control group before), that stayed untreated but were infected, the mortality was only slightly higher (36%). In contrast to these groups, the mortality was significantly reduced in groups where EPS was delivered into the water prior to and/or after the infection (2A and 4A with 4 and 7% mortality, respectively). While in group 2A with prophylactic EPS delivery only mild KHVD was observed, in group 4A the clinical signs were much stronger. The virus destroyed the entire skin to the extent that the fish skeletal tissue was exposed in affected fish.
The intensity of the clinical signs and the possibility to recover were estimated in the surviving fish at the end of the experiment in comparison to the untreated and non-infected negative control fish CB (Table 3). Visible signs of recovery like healed round or oval wounds in the skin ranged from 13% (2A) to 21% (3A) and were quite similar in range for all BM/EPS treated groups. Also the percentage of fish without any clinical signs in the groups with BM (prophylactic or metaphylactic) or prophylactic EPS delivery were almost the same, ranging from 78% (2A) to 84% (1A). A slightly lower percentage of fish with no clinical signs (69%) were registered for the 4A with metaphylactic delivery of EPS at the day of infection with KHV, where additionally 12% of the fish showed severe clinical signs of KHVD. In all other groups, no severe clinical signs like round or oval patches on the skin, fin rot or necrosis in gill tissues were visible during the external examination of the fish.
From the survivors of experiment 1, 10 fish were randomly chosen for serum collection from each group. Besides investigation by KHV antibody ELISA, all sera were also investigated by a newly established EPS antibody ELISA. All sera from negative control fish were negative by both antibody ELISAs. At least 40% of the sera had developed antibodies against EPS (Table 4). Interestingly, the response against EPS was slightly higher in fish treated with BM reaching 50 % in group 1A and 60 % in group 3A. Both prophylactically treated groups (1A BM and 2A EPS) developed a strong response against KHV with 90% and 70% responsive, respectively. In the metaphylactic groups 3A (BM) and 4A (EPS), only 40% and 20% had antibodies against KHV, respectively. Examination of the titres against EPS and KHV showed that both prophylactic groups, 1A (BM) with 1/76 and 2A (EPS) with 1/12, had much weaker titres against EPS and higher titres against KHV. In the metaphylactic groups 3A (BM) and 4A (EPS) it was the opposite: higher titres were obtained against EPS and weaker titres against KHV. When investigating the positive sera from the 10 fish chosen per group against EPS, only the sera from 1A showed an exceptionally high titre. All others were similar. Against KHV, the groups with BM delivery (1A and 3A) showed a weaker titres compared to the groups with EPS delivery (2A and 4A). The latter two groups showed an almost 50% stronger response against KHV based on the antibody titre.
Experiment 2 (EPS)
In the second experiment the influence of the various treatment set ups with EPS (prophylactic, metaphylactic or with increased concentration) were examined using only four groups of 50 fish. Group 1 and group 4 served as negative controls during the prophylactic six-week-delivery of EPS to group 2 (32 mg/mL) and group 3 (64 mg/mL). Within this time period, neither clinical signs nor any mortality were observed. After this period, all groups were challenged by immersion for 1 h with the isolate KHV-T. Afterwards, group 1 was used as a positive challenge control where no EPS treatment was applied. In the water of group 2, constant EPS delivery was continued for another 28 days. In group 3, EPS delivery was stopped and in group 4 EPS was delivered (32 mg/mL) after the challenge. Group 4 was used as metaphylactic EPS delivery group. At the end of the experiment no serological investigation was carried out.
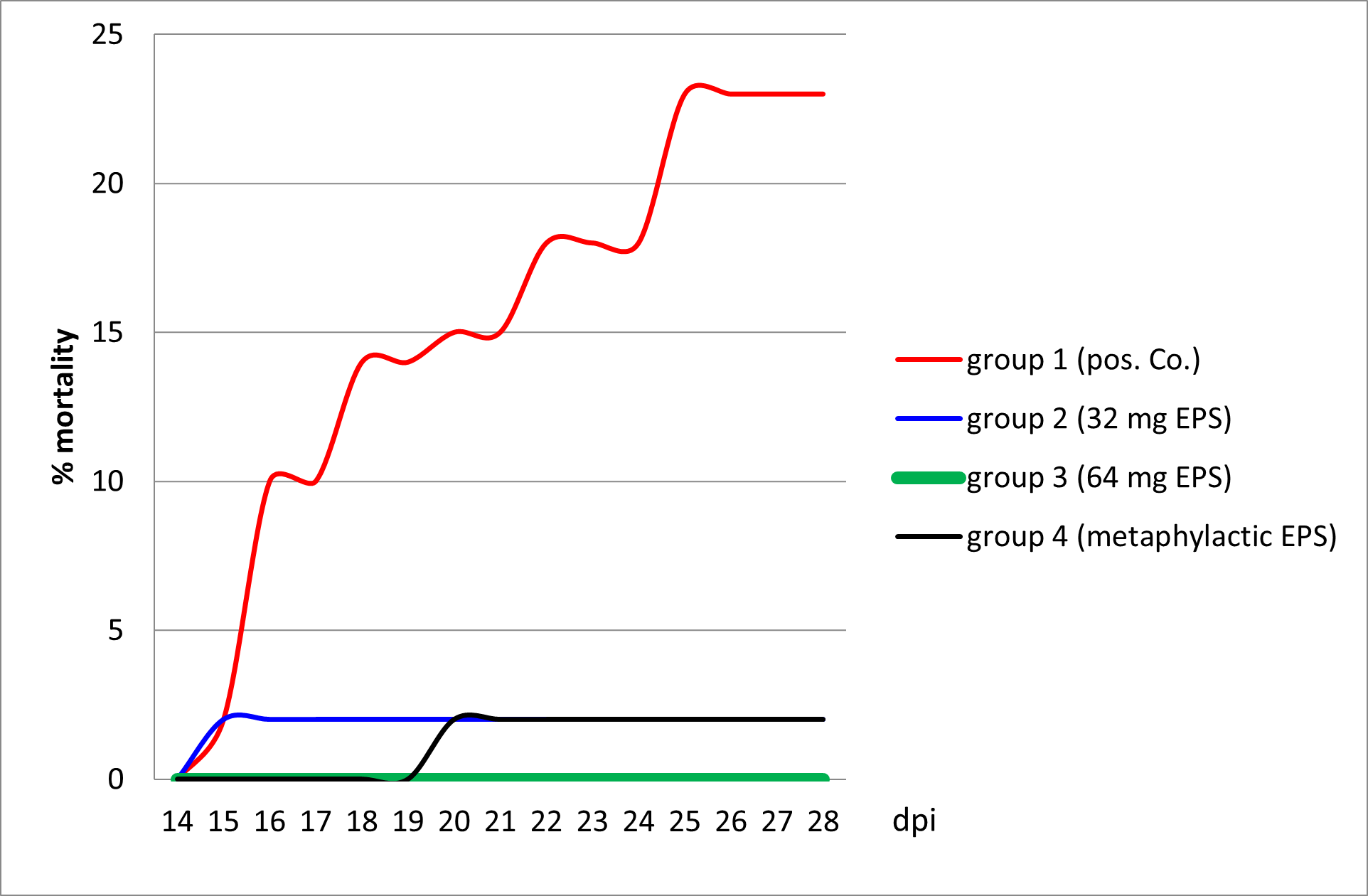
During this experiment, mortality in the positive control group 1 and in group 2 with 32 mg/mL delivery prior to, and after, the challenge started at 15 dpi. At 20 dpi, mortality started in group 4 that was treated with the same amount of EPS but only after the infection with KHV-T. No mortality at all was observed in group 3 that was treated prophylactically with doubled concertation of EPS before the challenge (Figure 1).

The influence of EPS delivery was observed after infection on the basis of the characteristics of KHVD development (Table 5). While in group 1 severe KHVD was accompanied by 23% mortality and 100% morbidity, in group 2 a weak mortality (2%) and 40% morbidity was observed with severe KHVD symptoms. In groups 3 and 4 very mild and mild KHVD was visible, but accompanied by a high morbidity rate (80 and 70%, respectively).
Discussion
Worldwide antiviral drugs which react against dangerous virus infections in all animals and in humans, without inducing adverse effects, have been under investigation for a long time (Bucknall 1973). Naturally occurring antiviral drugs, e.g. plants, mushrooms or parts of these, are the most interesting for use as therapeutics or metaphylactic measures (Lindequist et al., 2005). Often only one or two viruses are in the focus of the researchers (Reichert et al. 2017; McCaffrey et al. 2003). Research carried out by Yoshida et al. (2013) used bacteria from the gut of koi to influence KHV and assumed an antiviral effect of these bacteria. Other investigators showed that with an increasing number of bacteria which destroy cells, no cells are left for virus replication (van der Marel 2012). It was also suggested that high bacterial load might prevent virus entry to the host cell, thus causing an inhibition of replication (Kotob et al. 2016).
Further researchers used bacteria against viruses as a probiotic measure (Ibrahem 2015; Kamei et al. 1998; Li, Tan, and Mai 2009). While many bacteria in the gut or in the aquatic environment naturally produce EPS which can induce an inhibition of virus replication (Riccio et al. 2020), EPS from algae was successfully used to inhibit KHV replication and spread in in-vitro studies (Reichert 2016; Reichert et al. 2017). The advantage of EPS in low concentrations is that it does not damage the host or cells as bacteria can do. Additionally, EPS from different bacteria, often as an external component in the bacterial wall, seems to be effective against viruses, bacteria and fungi (Abdalla et al. 2021). Furthermore, ethanolic extracts from different microalgae were used to combat a human herpesvirus (Kok et al. 2011). EPS from microalgae is a waste product which occurs during the plateau phase of replication and consists, as other excretions, of polysaccharides, proteins, nucleic acids and lipids (Xiao and Zheng 2016). In this study, EPS was extracted from the culture supernatant of A. platensis and treated as described by Reichert et al. (2017). The basis for its use in-vivo was laid by the cell culture experiments with 32 mg/L which inhibited virus replication but had no negative effect on the cells, while EPS concentrations above 128 mg/L destroyed the cell monolayer depending on the cell culture. Our results shows that the cell lines CCB and EPC were more robust compared to E-11 cells when treated with EPS. Furthermore, the application of EPS prevented virus infection with all four investigated fish viruses, though at different effective concentrations.
To determine also whether the algae as a multifunctional organism could be used as a therapeutic measure, algae BM as well as excreted EPS were chosen to evaluate their in vivo reaction against KHV under wet facility conditions using carp as an experimental animal. For the prophylactic part of the experiments the BM and EPS content were counted according to daily and weekly water exchanges. In experiment 1, the BM content in the water was maintained constantly at 320 mg/L and of EPS at 32 mg/L. Additionally, in experiment 2 doubled EPS concentration was applied in water of one tank (64 mg/L). As described previously by others (Maizatul et al. 2017; Bergmann, Jin, et al. 2020) no influence on fish health and behaviour was observed due to the BM/EPS treatment during this period. Although a drop of oxygen content in the water was measured every time, two to three min after application for approximately 15 to 20 min, no damage to the fish was observed. Thus it is possible that the substances cover the oxygen probe leading to this short malfunction. After BM delivery, carp were covered in a green residue for two to three h. When EPS was delivered to the tanks, the water was transparent at all times and the fish were clearly visible. Both experiments were carried out at 18 to 20°C water temperature which is an ideal situation for carp and their immune reaction, but also for virus replication in the animals. For challenge with KHV, immersion was chosen to mimic the natural route of a waterborne infection. So far, there are no publications regarding use of EPS of A. platensis against KHV infection for comparison with the results obtained from this study. Regardless, other publications focused on alternative treatments, investigators had tried to cure KHVD by intracoelomically application of acyclovir with moderate success (Quijano Cardé et al. 2020).
In experiment 1, after infection with the European lineage virus KHV-E, a weak effect of BM against KHV was observed in the prophylactic and in the metaphylactic group. Nevertheless, the KHV induced mortality was almost the same as that in the positive untreated control group. In contrast, EPS applied in the prophylactic and metaphylactic (therapeutic) groups reduced the mortality significantly compared to all other groups. In the metaphylactic group 4A, the severe KHVD outbreak was prevented. In the prophylactic group 2A, KHVD broke out but with only weak clinical signs. The mortality rate at the end of experiment 1 was very similar in both these tanks (2A and 4A). In both groups, EPS obviously had an inhibitory effect on virus replication in the carp. Amazingly, the serological reactions against EPS and KHV differed considerably. While both prophylactic groups (1A and 2A) developed lower titres against EPS than against KHV despite a much longer delivery of BM and EPS, the opposite was observed in both metaphylactic groups (3A and 4A). This might be a sign of viral inhibition without a decreased humoral immune response in groups 1A and 2A, knowing that KHV is strongly immunogenic. On the other hand, in groups 3A and 4A virus entry may be blocked by BM and EPS, which at the end led to higher titres against EPS than against KHV because it was not necessary for the fish to develop antibodies to the latter. The average titres from the positive sera (out of 10) showed a uniform titre against EPS, except unexplainably the sera from group 1A. For so far unknown reasons, groups 1A and 3A developed weaker titres against KHV than the two EPS groups (2A and 4A).
To confirm the reaction of EPS against KHV, the second experiment was carried out with EPS only. Additionally, for challenge a highly virulent KHV-T isolate was used but at a lower temperature than usual for an Asian isolate. The temperature where KHV-T can develop its full virulent potential is between 23 and 28°C. In this study we tried to use European conditions also with an Asian KHV isolate and therefore performed both fish experiments at 20 °C. The development of the disease in experiment 2 showed a dependency of the survival rate from the EPS treatment. For unknown reasons the severity of KHVD was much stronger in group 2, which was treated with EPS before and after the challenge, but with a much lower morbidity rate compared to groups 3 and 4, that were either treated prophylactically with doubled EPS concentration or only metaphylactic with the same amount of EPS, respectively. While the morbidity rate was high in the latter two groups, the severity of KHVD was mild to very mild. Moreover, all EPS treated groups (2, 3 and 4) had a significantly higher survival rate compared to the untreated positive control group 1. The results confirmed the virus inhibiting effects of EPS against KHV in carp. This reaction may be associated with the inhibition of KHV replication but also with an increased immune response, perhaps as immunostimulant as assumed by Reichert et al. (2017).
Acknowledgements
The authors thank the team in Busan at FAU for the BM as well as the EPS. For her critical editorial review of the manuscript, the authors especially thank Mrs. Anette Beidler from FLI.