

Introduction
Streptococcal disease in tilapia (Oreochromis sp.) is a worldwide problem costing the cultured tilapia industry more than a billion dollars annually (Shoemaker, Xu, and Soto 2017). The most described and studied etiologic species are non-Lancefield groupable Streptococcus iniae (Shoemaker, Xu, and Soto 2017) and Group B S. agalactiae consisting of capsular types Ia, Ib and III (Shoemaker et al. 2017). However, additional species of streptococci are becoming pathogens of concern in wild and cultured fish populations. While originally believed to be restricted to saltwater fish hosts (Nomoto et al. 2006), Group C S. dysgalactiae is an emerging pathogen in both marine (e.g., Japanese yellowtail, Seriola quinqueradiata; amberjack, Seriola dumerili; pompano Trachinotus blochii) and freshwater (e.g., tilapia, O. niloticus; grass carp, Ctenopharyngodon idella; crucian carp, Carassius carassius) species (Costa et al. 2013; Hawke et al. 2021; Maekawa et al. 2020; Netto, Leal, and Figueiredo 2011)). Previous studies on the genetic diversity of geographically distinct alpha hemolytic S. dysgalactiae isolates from tilapia and other fish demonstrate these fish-associated strains form discrete clades, indicating host specificity (Abdelsalam, Eissa, and Chen 2015; Hawke et al. 2021).
Outbreaks of streptococcosis in fish may be complicated by concurrent infection of the host with other pathogenic microorganisms. Such co-infections in tilapia have been demonstrated to have negative impacts on health and performance (Abdel-Latif et al. 2020). Increased morbidity and mortality have been reported in co-occurring infections of Streptococcus sp. with parasites (Xu, Shoemaker, and LaFrentz 2014), viruses (Basri et al. 2020) and other species of bacteria (Sirimanapong et al. 2018). Concurrent infections may also result in immunosuppression (Assis et al. 2016) and can reduce vaccine effectiveness (Martins et al. 2011). In the present study, concurrent streptococcosis and columnaris disease was observed in Nile tilapia in the United States. Columnaris disease is another important disease impacting tilapia aquaculture, primarily caused by members of the family Flavobacteriaceae– Flavobacterium columnare, F. covae, F. davisii, and F. oreochromis (LaFrentz et al. 2022). Bacteria isolated from diseased tilapia were identified as Group C S. dysgalactiae subsp. equisimilis and F. davisii. The Group C S. dysgalactiae subsp. equisimilis isolates were confirmed and further characterised by phenotypic and genetic methods and an infectivity study used to fulfil Koch’s postulates.
Materials and methods
Collection of fish and bacterial isolation
A group of 900 Nile tilapia (mean length = 159 ± 8.1 cm and mean weight = 77.3 ± 9.4 g) were brought to USDA-ARS, Auburn, Alabama (16 January 2020) and placed in quarantine for overwintering. Following ~ 30 min of transport, fish were acclimated to 28°C and stocked into a 900 L tank maintained on flow-through dechlorinated city water. Initially the fish appeared normal. Eleven days after receipt, 1 dead fish was recovered and 4 fish with clinical signs of disease were observed. Clinical signs and gross changes included lethargy, frayed fins and discolored skin. Preliminary diagnosis following physical examination and skin scrape evaluation was columnaris disease, but with further examination, caudal peduncle pustules were also noted (Figure 1). Upon direct culture of the skin, brain, kidney and pustule on modified Shieh agar at 28°C, yellow pigmented, rhizoid, adherent colonies were cultured from the skin and kidney from one of 5 fish sampled. The same tissues cultured at 28°C on sheep blood agar (SBA) yielded gram-positive cocci from 3 out of 5 fish sampled. Culture of the pustules yielded pure gram-positive cocci (2 of 2 sampled). One yellow pigmented isolate (2K; recovered from the kidney of infected fish) and two gram-positive isolates (pustule-1 and kidney-2; recovered from the pustule and kidney, respectively) were further characterised. Overall mortality was low (<10 %). After diagnosis, fish were fed florfenicol (see below for susceptibility testing) -medicated feed (15 mg/kg body weight/day for 10 d) after which time no mortality was noted.

Identification and phenotypic characterisation of gram-positive isolates
The two gram-positive isolates (pustule-1 and kidney-2) were subjected to Lancefield (1938) grouping using Streptex™ rapid latex streptococcal grouping according to manufacturer’s instructions (Remel Europe Ltd, Kent, UK). Briefly, the isolates were grown on SBA plates for 24 h. Three to five isolated colonies were picked with a loop and subjected to the extraction procedure. The antigen extract (~20 µL) was mixed with a drop of the latex agglutination beads and observed for 1-2 min. Positive control multivalent antigen (provided with the kit) and negative control (no bacteria added) were included as controls.
The bacteria were incubated at 28°C for 24 h up to 4 d for observation of hemolysis on SBA and subjected to the API 20 strep strip to determine biochemical phenotype following the manufacturer’s protocol, with incubation at 30°C (bioMérieux Inc, Durham, NC, USA). We further evaluated the antimicrobial susceptibility of the isolates (pustule-1 and kidney-2) to oxytetracycline (T30), sulfamethazole trimethoprim (SXT) and florfenicol (FFC30) using the modified disk diffusion method (Bauer et al. 1966). Tests were completed at 28°C on cation adjusted Mueller Hinton agar (MHA) and 5 % SBA plates for recording the zone of inhibition after 24 and up to 72 h.
Identification of yellow-pigmented bacteria
The yellow pigmented bacteria exhibited rhizoid colony morphology and were adherent to the agar, consistent with the Flavobacterium spp. causing columnaris disease (LaFrentz et al. 2022). One isolate (2K) that was recovered from the kidney was identified to the species level using multiplex PCR (LaFrentz, García, and Shelley 2019). Briefly, gDNA (20 ng) from isolate 2K was used as template in the multiplex PCR, an appropriate positive control (consisting of a mixture of 10 ng gDNA each of F. columnare ATCC 23463T, F. covae AL-02-36T, F. davisii 90-106T, and F. oreochromis Costa Rica 04-02-TNT), and a no template negative control was included. PCR was performed as described by LaFrentz, García, and Shelley (2019). Following PCR, amplified products were resolved with 1.5% agarose gel electrophoreses and the size of the product was used for identification, with F. columnare, F. covae, F. davisii, and F. oreochromis yielding products with size of 415, 320, 894, and 659 bp, respectively.
Multilocus sequence typing of S. dysgalactiae
Multilocus sequence analysis (MLSA) of gram-positive isolates pustule-1 and kidney-2 was performed following the procedures adapted from Heckman et al. (2020). Isolates were revived from frozen stocks on SBA (48 h at 28°C) and individual colonies expanded in overnight cultures of brain heart infusion broth (Sigma-Aldrich, USA) at 28°C with shaking (150 rpm). Genomic DNA (gDNA) was isolated using the DNeasy Blood and Tissue kit (Qiagen, USA) following manufacturer’s recommendations for gram-positive bacteria. Quality and quantity of gDNA were assessed by a NanoDrop One Microvolume UV-Vis Spectrophotometer (Thermo Fischer Scientific, USA). PCR amplification of 9 housekeeping genes (arcC, glnA, groEL, gyrB, mutS, pheT, prkC, rpoB and tkt) was carried out using the primers from Heckman et al. (2020) with Phusion high-fidelity DNA polymerase (Thermo Fischer Scientific, USA) following manufacturer’s recommendations, with 3% added DMSO and annealing temperatures 6°C below the suggested melting temperature for each primer set.
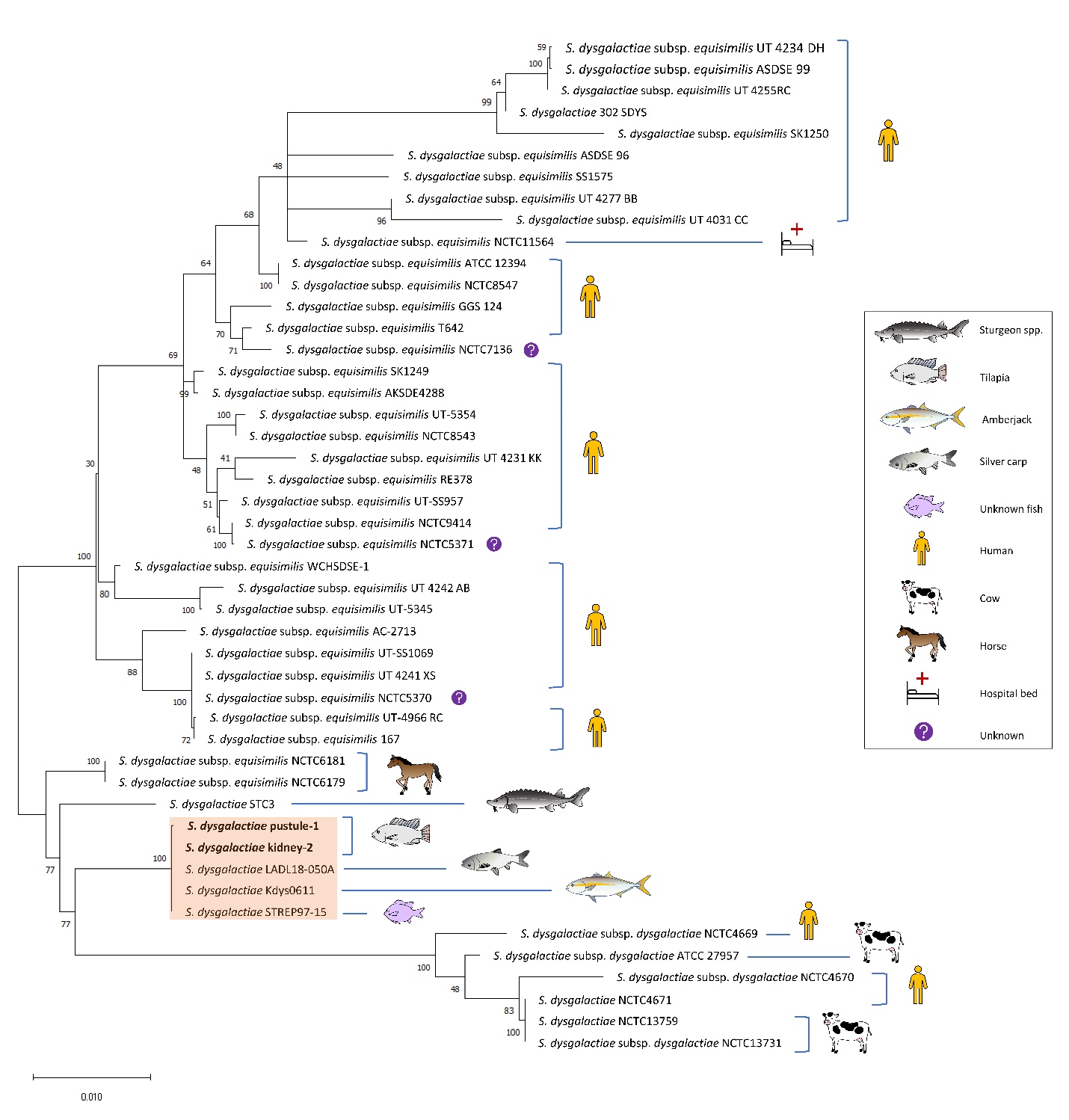
PCR products were electrophoresed to confirm fragment size, purified using the QIAquick PCR Purification Kit (Qiagen, USA) and assessed for concentration and quality. Purified products and corresponding forward primers were diluted and submitted for Sanger sequencing by GENEWIZ (San Francisco, CA, USA). Each housekeeping gene fragment was aligned by MUSCLE with default settings in Geneious Prime 2022.0.2 to corresponding sequences from the fish-associated S. dysgalactiae strain STC3 (Heckman et al. 2020; accession numbers MN955921.1, MN955970.1, MN956019.1, MN956068.1, MN956117.1, MN956166.1, MN956215.1, MN956264.1, and MN956313.1) and trimmed to the shared sequence length. A representative trimmed sequence of each housekeeping gene was used in BLAST searches of a localised database populated by the 44 S. dysgalactiae reference strains used previously for S. dysgalactiae MLSA by Hawke et al. (2021). The silver carp (Hypophthalmichthys molitrix) strain LADL18-050A was also included in analysis (Hawke et al. 2021). Sequences for each gene were concatenated alphabetically for each strain (arcC-glnA-groEL-gyrB-mutS-pheT-prkC-rpoB-tkt). The percentage of identical bases shared between the isolates and the reference strain sequences was determined by MUSCLE alignment of concatenated sequences to generate an identity matrix. For phylogenetic analysis, concatenated sequences were exported to MEGA-X (Kumar et al. 2018) and aligned by MUSCLE using the default settings. A maximum likelihood tree was generated using the Tamura three-parameter model with a gamma distribution assuming invariable sites (Tamura 1992), selected based on the Bayesian information criterion and the corrected Akaike’s information criterion in MEGA-X. The percentage bootstrap confidence levels were calculated from 1,000 resampling’s of the original data.
Streptococcus dysgalactiae infectivity study
Gift strain Nile tilapia (~40g) sourced from Florida (USA) were used to test the infectivity of the isolated S. dysgalactiae. All challenge procedures utilising fish were approved by the USDA-ARS, AAHRU Institutional Animal Care and Use Committee. Briefly, isolate pustule-1 was grown for 24 h at 28°C in tryptic soy broth (TSB) with shaking at 100 rpm prior to completing plate counts. Two tanks of fish (10 fish per tank) were injected intraperitoneally with 2.8 × 108 CFU/fish and a mock injected tank was injected with TSB only. The fish were housed in 57 L aquaria with flow-through water at 27 ± 2°C with dissolved oxygen ~ 6 mg/L. Fish were monitored for clinical signs of disease and mortality for 14 d. Direct inoculation (1µL disposable microbiological loop) of brain tissue from fresh dead fish were streaked onto SBA for re-isolation of the bacteria.
Results and discussion
Identification of Flavobacterium spp.
The isolate 2K was identified as F. davisii by the presence of an 894 bp product following multiplex PCR and agarose gel electrophoresis (Figure 2). This species is commonly isolated from tilapia with columnaris disease in the United States (B. LaFrentz, unpublished). Although an association between F. oreochromis and columnaris in tilapia has been demonstrated (LaFrentz et al. 2018), it is not surprising that the isolate was not F. oreochromis, as, to the authors knowledge, this species has not been isolated from fish in the United States.

Identification of gram-positive isolates and phenotypic characteristics
The isolated gram-positive bacteria were negative for hemolysis on SBA after 48 h and exhibited slight alpha (α) hemolysis after 3 to 4 d of incubation. Most S. dysgalactiae isolates described in the literature are α hemolytic on SBA (Nomoto et al. 2006). Results from the Lancefield grouping test demonstrated the gram-positive cocci isolated were Group C. Biochemical results yielded positive tests for ß- glucuronidase (ß-GUR), alkaline phosphatase (PAL), leucine aminopeptidase (LAP), arginine dihydrolase (ADH), acidification of ribose (RIB), acidification of trehalose (TRE), starch acidification (AMD) and acidification of glycogen (GLYG). The API 20 strep code was 0463013. This yielded a very good identification to the genus with the first choice of S. dysgalactiae, with the only test against being acidification of lactose (LAC). The S. dysgalactiae isolates described by Netto, Leal, and Figueiredo (2011) from Brazil were beta (ß) hemolytic and exhibited a similar phenotypic pattern except for variable results for hippuric acid (HIP), esculin (ESC), ADH and GLYG. Their (Netto, Leal, and Figueiredo 2011) isolates were also negative for LAC. The above phenotypes (i.e., grouping and biochemical reaction) suggested our isolates were Group C S. dysgalactiae.
Multilocus sequence analysis
MLSA confirmed the identification of the isolates as S. dysgalactiae and both isolates grouped with the other strains isolated from fish (Figure 3). Pustule-1 and kidney-2 shared 99.9-100% concatenated sequence identity to S. dysgalactiae strains LADL18-050A from carp in the USA (H. molitrix), Kdys0611 from amberjack in Japan (Seriola dumerili; Nishiki, Yoshida, and Fujiwara 2019) and STREP97-15 from an unspecified fish species in Singapore (Koh, Binte Abdul Rahman, and Sessions 2020). Strain STC3 from sturgeon in Uruguay (Acipenser sp., Heckman et al. 2020) was most divergent from the other fish isolates, with only 98.6% shared sequence identity. The fish isolates grouped most closely to S. dysgalactiae subsp. equisimilis strains from horses, and shared overall higher sequence identity with all S. dysgalactiae subsp. equisimilis strains than with those in subspecies dysgalactiae. This is in line with recent findings by Porcellato et al. (2021), demonstrating that while previous studies assigned fish-associated S. dysgalactiae strains to subsp. dysgalactiae, they are actually more related to other strains of S. dysgalactiae subsp. equisimilis isolated from animals. The MLSA also supports the existence of fish-specific clades within the subspecies (Porcellato et al. 2021; Hawke et al. 2021). Further investigation, including incorporation of additional isolates into MLSA and virulence testing, complemented with whole genome phylogenetic analysis will better define these genetic groups and their importance in fish disease.

Antibiotic susceptibility/resistance
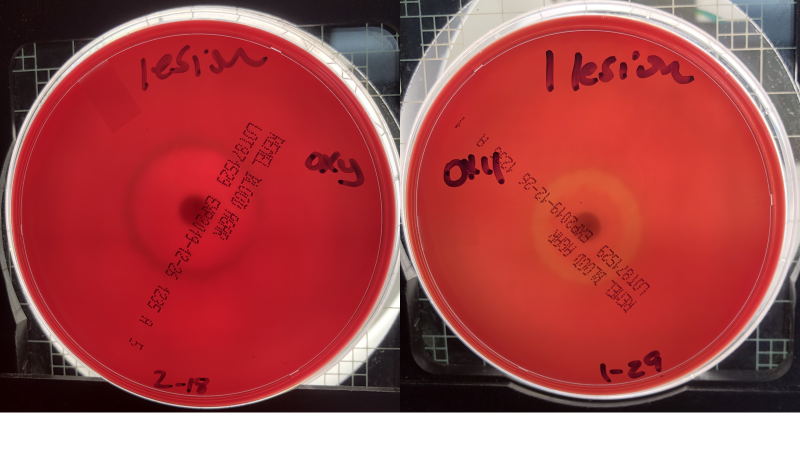
Both S. dysgalactiae subsp. equisimilis isolates yielded susceptible phenotypes (Table 1) based on susceptible zone diameter interpretive standards of > 19 mm SXT, > 17 for FFC30, > 26 for T30 (NCCLS 1994), although specific cut offs have not been established for S. dysgalactiae subsp. equisimilis. We initially tested antibiotic sensitivity on 5% SBA plates and then on the suggested medium (cation adjusted Mueller Hinton Agar) used for antibiotic testing. On both media, the zone of inhibition was mostly similar. Interestingly, on SBA, we noted the induction of hemolysis at or near the zone of inhibition with two of the antibiotics tested (oxytetracycline and florfenicol) with α hemolysis seen at 24 h and ß hemolysis noted by 72 h (Table 1 and Figure 4). Upon a literature search, we noted one paper on Propionibacterium acnes, a non-spore forming, anaerobic, gram-positive organism, responsible for invasive infections post shoulder surgery (Wright et al. 2016). They (Wright et al. 2016) noted the induction of hemolysis upon exposure to antibiotics on SBA as well, and suggested hemolysis induction was related to clindamycin resistance. In our case, although we only tested two isolates and the approved antibiotics for fish, hemolysis induction did not appear related to zone of inhibition. The phenomenon observed in our case may be related to a bacterial stress response post antibiotic exposure upregulating hemolysin production. No increase in mortality was observed after the fish were fed florfenicol for the required duration, so in vivo hemolysis induction may not have occurred. Fish showing pustules did not appear to recover and those animals with severe pustules were euthanised. More studies are needed to assess the induction of hemolysis in vivo following feeding antibiotics and or in vitro using whole fish blood to establish biological relevance.
_showing_alpha_(.png)
Infectivity results
No mortality was observed in the mock infected fish. The infected fish yielded 10% (1/10) and 20% (2/10) mortality in the replicate tanks. Although the mortality was low, Koch’s postulates were fulfilled. Clinical signs included lip, caudal and dorsal fin pustules (data not shown). Streptococcus dysgalactiae subsp. equisimilis was recovered on SBA from the dead fish (3/3). Based on the dose of bacteria injected, we expected to see mortality near 100% as observed by Netto, Leal, and Figueiredo (2011). Interestingly, the mortality obtained in our infectivity study was low grade, like the infection noted in the case study, although the case study fish were treated with antibiotics. The differences between our study and the study published by Netto, Leal, and Figueiredo (2011) were fish size (40 g vs 120 g), water temperature (28 vs 32°C) and hemolysis (alpha vs beta). Any or all factors may have contributed to the differences noted.
Coinfections of tilapia with multiple pathogens are reported in the literature (Abdel-Latif et al. 2020). Interestingly, this appears to be the first case of S. dysgalactiae susp. equisimilis and F. davisii co-occuring. We did not test F. davisii infectivity in this study as research describing the virulence of columnaris causing bacteria in tilapia is well known (Shoemaker and LaFrentz 2014). The tilapia in this study likely carried the pathogens and the transport stress triggered the low-grade mortality and clinical signs noted. The work highlights the impact of concurrent infections on tilapia and the potential to unknowingly move multiple pathogens during transport.
Acknowledgements
The authors acknowledge the excellent technical assistance of Mrs. Paige Mumma (USDA-ARS). This research was supported by USDA-ARS CRIS Project No. 6010-32000-027-00-D, Integrated Research to Improve Aquatic Animal Health in Warmwater Aquaculture. Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the United States Department of Agriculture.