__a_potential_clinical_sign_of_in.tiff)


INTRODUCTION
Lumpfish (Cyclopterus lumpus) are widely used commercially as cleaner fish to delouse farmed Atlantic salmon (Salmo salar) of Lepeophtheirus salmonis. The aquaculture industry is largely dependent on both wild caught and farmed lumpfish for their supply (Imsland et al. 2014) and little is known about the diseases that may adversely affect them. Cyclopterus lumpus virus (CLuV) is a flavivirus and was first reported in lumpfish in 2017 (Skoge et al. 2017). An additional flavivirus was later identified affecting chinook salmon (Oncorhynchus tshawytscha) (Soto et al. 2020). Flaviviruses are members of the Flaviviridae family, which are single stranded, positive-sense RNA viruses with genomes of approximately 9 – 13 kb (Simmonds et al. 2017). CLuV affects the liver, previously reported to cause hepatocellular degeneration and lipid accumulation leading to an increase in mortality (Skoge et al. 2017). Whilst little is known about the full impact of CLuV on both farmed and wild lumpfish, less is known about whether lumpfish can act as vectors for salmon diseases of major concern, since some pathogens possess a broad host species range. For example, both viral haemorrhagic septicaemia virus (VHSV) and Renibacterium salmoninarum (the causative agent of bacterial kidney disease) have been previously reported affecting lumpfish (Guðmundsdóttir et al. 2018; Gnanagobal et al. 2021). Therefore, lumpfish could act as a vector for the transmission of these, and indeed other significant pathogens of Atlantic salmon.
In September 2021 a site in Dorset, England, reported increased mortalities of lumpfish stocks ranging from 1-3 % daily, up to 30 % mortality in total, instigating a disease investigation by a local veterinarian and Cefas Fish Health Inspectorate (FHI).
This study describes the results of comprehensive disease investigation using isolation of viruses on cell culture, bacteriology, histopathology, and molecular diagnostics. Since information on the pathogen susceptibility of lumpfish is still limited, our broad approach was to rule out possible pathogens that may not only affect the lumpfish themselves, but also the salmon kept in close contact.
MATERIALS AND METHODS
A gross assessment of any changes to the skin, mucous, gills and internal organs for clinical signs of disease was carried out by a MOWI veterinarian. Tissues were also collected for commercial reverse-transcription real-time polymerase chain reaction (RT-rPCR) testing.
In the follow up site visit by the Cefas FHI, a total of 40 fish were sampled across 3 tanks. Pools of 10 fish per pool were prepared for laboratory analysis. The liver, kidney, spleen, heart, gill, and gastrointestinal tract (GIT) were sampled for histology and placed into 10 % neutral buffered formalin (NBF) for a minimum of 24 h fixation. Tissues were subsequently processed for formalin fixed paraffin embedded (FFPE) histology using a Thermo Excelsior AS (Thermo scientific, UK) vacuum infiltration processor following routine protocols. Tissue sections of 3-5 µm thickness were obtained using a Leica histocore multicut microtome (Leica biosystems, UK) and stained with haematoxylin and eosin (H&E). Slides were examined and pictures were captured using a Nikon Ni-E brightfield microscope and Nikon DS-Ri2 digital camera (Nikon, UK) respectively.
For molecular screening and virus culture, samples of kidney, spleen, brain, and liver (to a total weight of approximately 1 g per fish pool) were placed into virological transport medium (Glasgow MEM (Life Technologies, UK) that was supplemented with 10 % foetal bovine serum (Merck. UK), GlutaMAX (Life Technologies, UK), and 1 % antibiotic and antimycotic solution (Merck, UK). All samples were placed into chilled cool boxes and transported back to the laboratory within 48 h of the sample time. The tissues were removed from the transport medium, homogenized with a pestle, mortar and sterile sand, then re-suspended in the original transport medium and centrifuged at 3100 RPM for 15 min at 4 °C.
For the molecular screening, total nucleic acid was extracted from 100 µL aliquots of the clarified tissue homogenate using the Virus Mini Kit and EZ1 extraction robot (Qiagen, UK) using the manufacturers recommended protocols, and the nucleic acid was eluted in a 60 µL volume. These total nucleic acid extractions were tested for infectious salmon anaemia virus (ISA), ranavirus, salmon alphavirus (SAV), and CLuV, as part of the complete disease screen. The ISA and ranavirus assays were carried out following the RT-PCR and PCR protocols stipulated by the world organisation for animal health (WOAH – previously OIE) aquatic manual, (WOAH 2022). Samples were screened for SAV using the RT-PCR method described by Cano et al. (2014), and CLuV using the RT-rPCR described by Skoge et al. (2017). The reverse transcription was performed in a 20 µL volume containing 4 µL MMLV 5X RT buffer, 1 µL dNTPs, 1 µL RNasin, 1 µL random primers, 1 µL MMLV RT enzyme (Promega, UK) and 4 µL RNA template, made to 20 µL with molecular grade water. The real-time assays were performed in a 20 µL reaction volume containing 10 µL TaqMan universal master mix (Life technologies, UK), 5.1 µL molecular grade water, 0.8 µL primer and probes (Table 1) and 2.5 µL cDNA template. Assays were performed on a Quantstudios 3 thermocycler (Applied Biosystems. UK) using cycling settings with an initial hold stage of 95 °C for 10 min, followed by 50 cycles of 95 °C for 15 sec, and 60 °C for 1 min. To confirm any real-time positive results for CLuV, a set of conventional primers were designed (Table 1) based on the published data for the CLuV NS5 polymerase region of the polyprotein gene (Accession NC_040555.1).
The remainder of the cDNA generated for the RT-rPCR was used for the conventional RT-PCR assays. The PCR reactions were performed in a 50 µL volume containing 2.5 µL cDNA samples in reaction volumes of; 0.5 µL of 25 mM dNTPs, 10 µL 5x green GoTaq flexi buffer, 5 µL of 25 mM MgCl2, 0.25 µL GoTaq G2 Flexi DNA Polymerase (Promega, UK), 0.5 µL of forward and reverse primer at 10 pmol, and 30.75 µL of molecular grade water. The cycling program had an initial denaturation at 94 °C for 5 min, followed by 40 cycles of 94 °C for 1 min, 55 °C for 1 min and 72 °C for 1 min, with a final extension of 72 °C for 10 min. PCR products were resolved on a 2 % agarose gel (w/v) agarose/TAE (40 mM Tris-acetate, pH7.2, 1 mM EDTA) containing 1.0 µg/mL ethidium bromide, at 120 V for 30 min. Products of the expected size were excised, and the DNA purified using the freeze and squeeze method using spin modules (MP biomedicals, UK). Both strands of the DNA were sequenced by Sanger sequencing using big dye terminator 3.1 methodology (Life technologies, UK) using the manufacturer’s recommended protocol and the sequences were analysed on a 3500xl genetic analyser (Applied Biosystems, UK). Consensus sequences were generated using CLC workbench 7 software (Qiagen, UK)
For the isolation of virus on cell culture, the clarified supernatant was passed through 0.45 µm sterile filters (Sartorius, UK), then inoculated onto bluegill fry (BF-2), Epithelioma papulosum cyprini (EPC), Chinook salmon embryo (CHSE-214), and E-11 (clone of SSN-1), all cell lines taken from the WOAH – (previously OIE) Manual of Diagnostic Tests for Aquatic Animals, (WOAH 2022). Cell lines were inoculated on falcon 12 well culture plates (Scientific Laboratory Supplies, UK), and incubated at 15 °C, 20 °C and 25 °C, for 14 days , including a passage on day 7. Tests were undertaken for viral haemorrhagic septicaemia virus (VHSV), infectious haematopoietic necrosis virus (IHNV), infectious pancreatic necrosis virus (IPNV), nervous necrosis virus (NNV) and CLuV, via observations of cytopathic effect (CPE) .
Bacterial swabs (transwabs) containing AMIES transport medium (VWR, UK), were taken for bacteriology. These swabs were sub-cultured on to sea-water agar (SWA, Oxoid, UK) with plates and incubated at 22 °C. These plates were observed every 18-24 h for significant growth and/or morphological appearances. Sub-cultures of those colonies were performed to ensure purity suitable for culture characterisation using preliminary identification tests (e.g., Gram staining, motility, catalase, oxidase, and oxidative/fermentation glucose testing). A specific analytical profile index (API, BioMerieux, France) test was also used on these colonies to record enzymatic activity. This data was added to the bioMerieux API profiling database. For confirmation, colonies were collected and placed into 100 µL of molecular grade water and tested using 16S rRNA PCR incorporating fD1 and rP2 primers (Weisburg et al. 1991), and GoTaq G2 Flexi DNA Polymerase (Promega, UK), for 30 cycles followed by Sanger sequencing method as described above.
RESULTS
The moribund fish originally examined by the veterinarian appeared to show disease signs including increased mortalities, and pre-nursery fish were lethargic, attaching to the side or bottom of tanks, and showed a loss of appetite. Similar behaviour was observed in nursery fish that had been transferred from the same batch during the previous week. Changes observed by an autopsy suggested an infection by lumpfish flavivirus (CLuV) (Figure 1), and the subsequent commercial RT-rPCR test provided a positive result for CLuV.
__a_potential_clinical_sign_of_in.tiff)
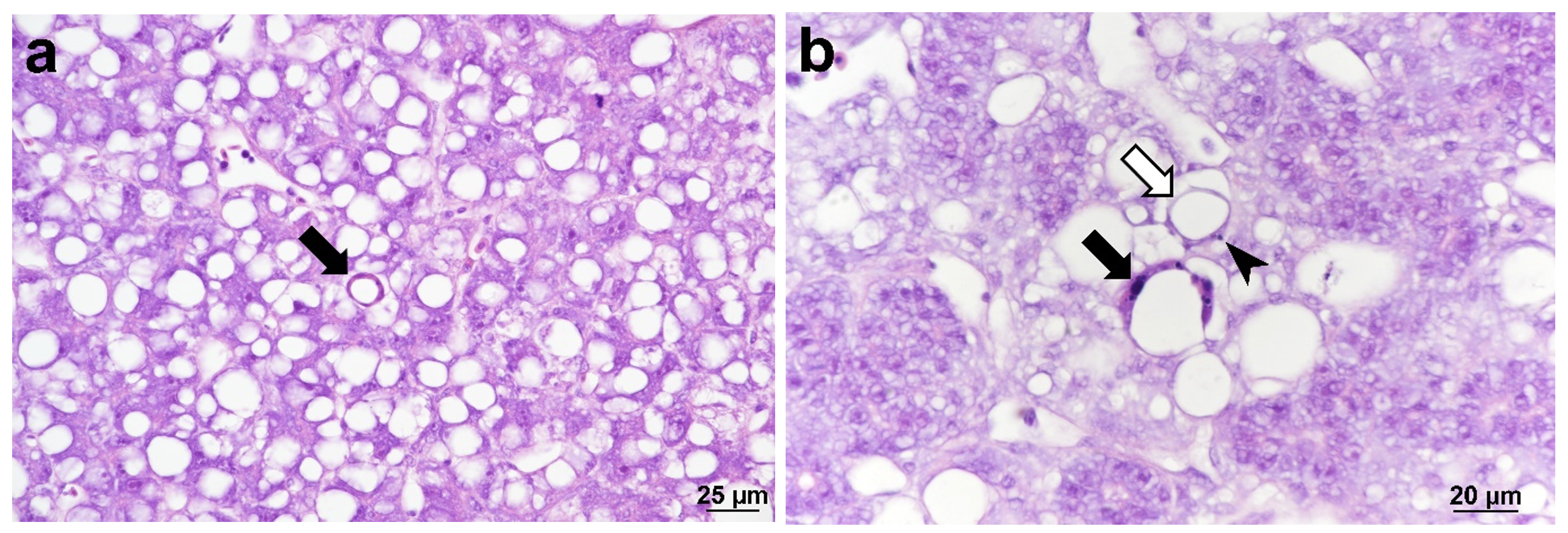
The Cefas investigation confirmed the presence of CLuV using histopathological and molecular techniques. Histological examination primarily revealed mild multifocal hepatocellular degeneration that generally appeared characteristic of programmed cell death (apoptosis), characterised by cellular shrinkage and karyorrhexis, albeit with an atypical presentation (Figure 2). Cell death appeared to be responsible for the formation of large vacuoles within the parenchyma. However, varying degrees of macrovesicular steatosis were also observed within samples, characterised by a single large vacuole within hepatocytes resulting in nuclei and other organelles being displaced towards the cell periphery. Considering this, the atypical apoptotic presentation may result from vacuolated apoptotic hepatocytes shrinking tightly around these large lipid vacuoles.

The RT-rPCR assay for CLuV was positive, with Ct values of approximately 19 to 29 for all 4 samples (Figure 3). Similarly, products with the expected size of 568 base pairs (bp) were also obtained for all samples using conventional PCR assay (Figure 3).

Sequence analysis confirmed that PCR products, shared 99.63 % nucleotide identity homology to one entry for lumpfish flavivirus (NC_040555.1) with two synonymous nucleotide substitutions at position 10112 and 10136 in the genome being the only major difference., This new sequence was confirmed as CLuV and has been assigned the accession number ON287380, and 99.44 % to the only other entry (MF776369.1) (Figure 4).

The ISAV, SAV and ranavirus specific assays were negative (data not shown).
For virology, no cytopathic effects were observed in the BF, EPC, CHSE-214 or E-11 cell lines at any of the temperatures used.
Bacteriology and 16S rRNA analysis showed no significant colonies but did reveal some environmental bacteria that are usually found in and around arctic waters, including Lacinutrix sp (MW580188.1) and Psychromonas arctica (AJ308372.1).
DISCUSSION
We discovered a CLuV infection in lumpfish on a farm site in the south of England in September 2021, which is the first time on record in England. The fish were showing the typical clinical signs of CLuV and were positive following an initial screen with the published RT-rPCR assay (Skoge et al. 2017). We also applied histopathological methods and developed a conventional PCR assay to confirm this finding. Unfortunately, we were unable to cultivate the virus on the cell lines we had available, which matched observations by Skoge et al. (2017)
It is worth noting that although likelihood of bacterial infection with known salmonid pathogens is low (Johansen et al. 2016) there was the possibility of infection via an atypical Aeromonas salmonicida strain (VKM et al., 2017). However, in this study, bacteriological isolation did not provide evidence for any significant infections, only environmental strains were present. Whilst Atlantic salmon were generally considered to be non-susceptible to VHSV, more recently both the salmon and lumpfish were shown to be infected with a novel VHS strain in genotype IV (Erkinharju et al. 2021). Atlantic salmon were also shown to be susceptible to NNV in challenge studies (Korsnes et al. 2005), but in previous studies, infected lumpfish showed no signs of histopathology or nervous behaviour, suggesting the virus in lumpfish may be undetectable but may be transmissible (Toffan et al. 2019) reinforcing the concern that lumpfish could potentially be a vector for disease, including CLuV. Other salmonid viruses, such as SAV or ISA, have not yet been detected in lumpfish, but they have been reported in other wild fish vectors such as herring (Nylund et al. 2002), wrasse (Røsaeg et al. 2017), and flatfish (McCleary et al. 2014). In this study we did not detect CLuV by culture and failed to detect any evidence of any other RNA or DNA viruses by RT-PCR and PCR respectively.
Currently there is no information on the prevalence of CLuV in the wild lumpfish populations globally, and with no molecular epidemiological data from Norway (Sommerset et al. 2021), it is difficult to ascertain if the virus had been introduced to the UK farm site via imported juveniles or transmitted to the farmed fish from a local source of the virus. It is also worth noting, that in the absence of epidemiolocal data, it remains a possibility that the virus is endemic and that stressors caused the sudden increase in mortalities. Due to SARS-Cov-2 restrictions in place at the time, stocks of fish remained unsold and had grown larger than usual, this overcrowding caused an increase in aggression within the lumpfish population. In addition, at the time of the mortality, problems with a blocked filter may have caused an increase in dissolved organic solids phosphate (PO3/PO3) and presence of a low concentration of hydrogen sulphide (H2S) and/or bromine. Once the site identified and rectified these issues, the mortalities reduced significantly and returned to the normal levels.
Without a better understanding of the host range, or the epidemiology of CLuV, any potential long terms risks to the salmon industry and impacts on wild fish may go unchecked. Therefore, in future studies it will be essential to consider the best approaches to undertake the experiment challenge work required to fully evaluate these risks.