
Introduction
Viral nervous necrosis (VNN) is a significant viral disease caused by betanodaviruses, particularly in larvae and juveniles and rarely in adults. VNN is depicted as one of the most economically threatening problems for marine aquaculture (Fernández Sánchez et al. 2022). With the industry beginning to culture different fish species, VNN outbreaks cause concerns about sustainability (Costa and Thompson 2016; Thiéry et al. 2011; Panzarin et al. 2010; Toffan et al. 2017).
Betanodaviruses are classified in the Nodaviridae family and have an icosahedral structure and a round morphology, are non-enveloped with a diameter of approximately 25 nm (Breuil et al. 1991). Their genomes are single-stranded with positive polarity and two segments, RNA1 and RNA2 (Johnson, Price, and Ball 2003). The RNA1 (3.1 kb) is responsible for the synthesis of RNA-dependent RNA polymerase (RdRp) and is involved in the adaptation of the virus to temperature (Ahlquist et al. 2003; Gallagher and Rueckert 1988; Hata et al. 2010; Nagai and Nishizawa 1999; Panzarin et al. 2014). During the replication of RNA1, subgenomic RNA3 transcript production is directed by RdRp and contributes to the replication process (Fenner, Goh, and Kwang 2006; Iwamoto et al. 2005; Johnson, Price, and Ball 2003). The RNA2 (1.4 kb) segment encodes the capsid protein and is responsible for host tropism and immunoreactivity of different genotypes (Ito et al. 2008; Iwamoto et al. 2004; Johnson, Price, and Ball 2003; Nishizawa et al. 1995; Panzarin et al. 2016).
According to phylogenetic analysis of the T4 variable region of the RNA2 segment, betanodaviruses are grouped into four major genotypes, including striped jack nervous necrosis virus (SJNNV), tiger puffer nervous necrosis virus (TPNNV), barfin flounder nervous necrosis virus (BFNNV), and red-spotted grouper nervous necrosis virus (RGNNV) (ICTV 2021). Rather than being species-specific, genotypes require a water temperature at which they develop optimally (Costa and Thompson 2016; Mori et al. 2005). Because of the segmented nature of the virus, the formation of reassortant strains has thus far been identified in the SJNNV and RGNNV genotypes, and RGNNV/SJNNV or SJNNV/RGNNV reassortants have been reported many times in sea bass, sea bream and sole (Bitchava et al. 2019; He and Teng 2015; Kaplan, Pekmez, Cagirgan, et al. 2022; Toffan et al. 2017; Volpe et al. 2020).
Betanodaviruses are commonly referred to as nervous necrosis viruses (NNV). So far, at least 173 different cultured or wild fish and invertebrate species have been susceptible to NNV and the virus has caused epidemics in at least 62 of them (Bandín and Souto 2020). Worse still, this number increases day by day. NNV typically causes outbreaks in farmed fish, but severe outbreaks have also appeared in wild fish, particularly grouper (Gomez et al. 2009; Valencia et al. 2019; Vendramin et al. 2013). Asymptomatic NNV infections have also been detected in wild fish (Baeck et al. 2007; Barker et al. 2002; Ciulli et al. 2007; Gomez et al. 2004, 2008b; Liu et al. 2014; Panzarin et al. 2012). Bivalve mollusks or crustaceans can also play a role in the spread of various NNV strains (Berzak et al. 2019; Ciulli et al. 2010; Gomez et al. 2008a; Kim et al. 2017; Panzarin et al. 2012).
In recent years, research on different geographical regions has shown that RGNNV is common not only in farms but also among wild fish in the Mediterranean basin and on the coasts of Asia and Australia (Bandín and Souto 2020). The lack of effective treatment for the disease and the introduction of applied management such as monitoring the spread of the virus helps to reduce the effects of the disease (Doan et al. 2016). For this reason, surveys of farm and wild fish in different regions provides important information, increasing the capacity for early warning systems and awareness, and beyond that increasing biosecurity, including implementation of vaccination regimes. This study aimed to investigate the presence of betanodavirus in wild fish along the Turkish Mediterranean coast, where betanodaviruses are endemic, and to characterize those circulating isolates.
Materials and methods
Sampling
Along the Mediterranean coast of Turkey, 400 wild fish belonging to 27 species were caught by fishing between 2019 and 2021 (Table 1). Samples were collected from five different regions of the Mediterranean coast of Turkey, which were Antalya Bay, Mersin Bay, Iskenderun Bay, and Finike and Manavgat coasts (Figure 1).

Preparation of samples
The fish, transported to the laboratory through the cold chain, were examined macroscopically on an individual basis. Brain, eye and spinal cords were removed following laboratory biosafety rules. The organs were homogenized (1/5, w/v) in a refrigerated homogenizer (Allsheng/BIOPREP-24) with ceramic beads in EMEM (Eagle’s Minimum Essential Medium) (Sigma‒Aldrich, United Kingdom). The EMEM contained 2% fetal bovine serum (FBS) and 2% antibiotics (Sigma, USA). The homogenates were centrifuged at 4℃ and 4000 × g for 15 min (ThermoFisher SL 16R, Germany). Supernatants were collected and stored at -80℃ until use.
Total nucleic acid extraction
RNA extraction was done in an automatic extraction device (Roche MagNA Pure LC System, Germany) following the protocol of the commercial kit (MagNA Pure LC Total Nucleic Acid Isolation Kit, Roche, Germany) using 200 µL from each supernatant. Positive (TR.VNN.01.01) and negative (EMEM) controls were used to validate the extraction process and check cross-contamination.
Real-Time RT-PCR
Real-time reverse transcription polymerase chain reaction (RT-rPCR) was performed to detect the presence of NNV. Primers and probes specific to the T4 variable region of the RNA2 segment of betanodavirus (Panzarin et al. 2010) were used. RT‒rPCR was performed using a commercial kit (Real Time Ready Virus Master, Roche, Germany) and real-time PCR device (Roche LightCycler® 480 II). Mastermix was prepared as 7.6 µL H2O, 1 µL F primer (10 µM), 1 µL R primer (10 µM), 1 µL Probe (10 µM), 4 µL 5x buffer and 0.4 µL enzyme mix for each sample. Then, 15 µL of each was added to 5 µL of template and total volume was 20 µL. Positive and negative controls were added to each reaction to verify the PCR process and a contamination control. After being covered with transparent tape, the plate was centrifuged at 1500 g at 4℃ for 2 min and placed in the PCR device. Reaction conditions were set as 50℃ for 10 min for reverse transcription, 95℃ for 65 sec for the inhibition of reverse transcriptase, 95℃ for 10 sec for pre- denaturation, 54℃ for 30 sec for annealing, and 72℃ for one sec for 45 cycles for elongation. The study followed the previously optimized RT‒rPCR method (Kaplan and Karaoğlu 2021).
cDNA synthesis and PCR
cDNA synthesis (EvoScript Universal cDNA Master, Roche, Germany) was performed from the sample extracts to store the positive samples for longer periods of time and to perform sequence analysis. For cDNA synthesis, the mixture consisted of 4 µL of reaction buffer (5x conc.), 9 µL of H2O and 5 µL of sample RNA. Reaction conditions were as follows: 42℃ for 15 min, 85℃ for 5 min, 65℃ for 15 min, and 4℃ for post-reaction standby.
PCR was performed (Techne TC-412, United Kingdom) using a commercial kit (Xpert Fast Hotstart Mastermix with Dye, GRISP, Portugal) and specific primers (Bovo et al. 2011; Toffolo et al. 2007) from positive sample cDNAs were used to obtain partial genome sequences of both RNA1 and RNA2 segments of betanodavirus. The total reaction mixture consisted of 25 µL with: 12.5 µL master mix, 5.5 µL water, 1 µL forward (10 mM) and 1 µL reverse primers (10 mM), and 5 µL cDNA for each sample. The amplification cycle was 95℃ for 3 min of pre- denaturation followed by 45 cycles at 95℃ for 15 sec of denaturation, 62℃ for 15 sec of annealing, 72℃ for 15 sec of elongation, and one cycle for 3 min at 72℃ for the final elongation.
The amplicons were treated with DNA loading dye (6x Loading Dye, Thermo Scientific, USA, R0611) and transferred to a 1.5% agarose gel (Sigma, USA) containing 0.5 µg/mL ethidium bromide (Nalgene Fisher Scientific, BP-1302-10, USA). A 100 bp marker (GeneRuler 100 bp DNA Ladder, Thermo Fisher Scientific, Lithuania, SM0241COMPONENTA) was also used to determine the product size. Electrophoresis was set at 80 volts for 60 min. Then, the agarose gel was examined on an imaging device (Vilber Lourmat, EEC, France).
Sequencing and phylogenetic analysis
Positive samples were submitted to partial genome sequencing for both RNA1 and RNA2 segments of the virus, performed by a commercial company (Microsynt, Balgach, Switzerland) to determine the genotype and to perform phylogenetic analysis. Correction and matching of the results were performed with the DNADynamo (Blue Tractor Software Ltd., UK) software. The resulting consensus nt sequences were compared and confirmed in the Basic Local Alignment Search Tool (BLAST) system of the National Center for Biotechnology Information (NCBI) (Altschul et al. 1990). Subsequently, accessible close isolates in GenBank and previously detected isolates in the Mediterranean were included in the molecular study, except for very short sequences. All sequences to be studied were aligned with the ClustalW method using the MEGAX software (Kumar et al. 2018). Phylogenetic analyses were performed by determining the best protein/DNA model for both segments. The nt and amino acid (aa) similarity rates between the sequences were determined according to both RNA1 and RNA2 segments. Using the maximum likelihood (ML) method, 1000 replicate bootstraps were selected, and phylogenetic trees were created (Kumar et al. 2018).
Results
During the necropsy in the laboratory, macroscopic examinations did not reveal any lesions in the fish.
The NNV genome was detected in two out of four garfish (Belone belone) sampled in Iskenderun Bay (Figure 1) during October 2019. Both positive samples were collected in the same region and one used for further studies was named as TR. VNN.01.03. NNV was not detected in other samples neither in 2019 nor in the samples from March to April in 2020 and 2021. In addition NNV was detected in high prevalence in red mullet (Mullus barbatus) collected in the Antalya Gulf (25/50) in September 2021. One of them was kept for further molecular studies and the isolate was named as TR.VNN.07.01.
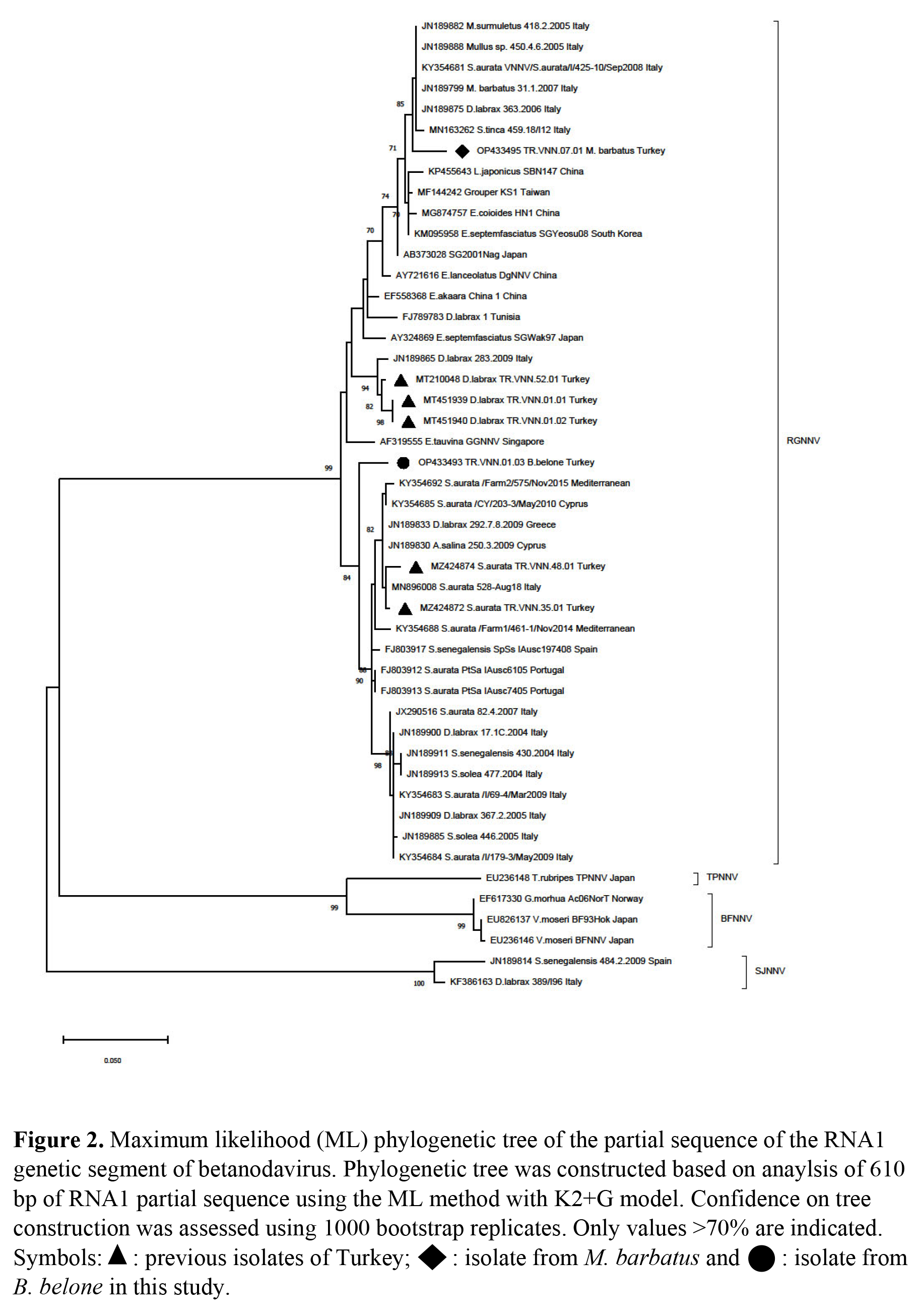
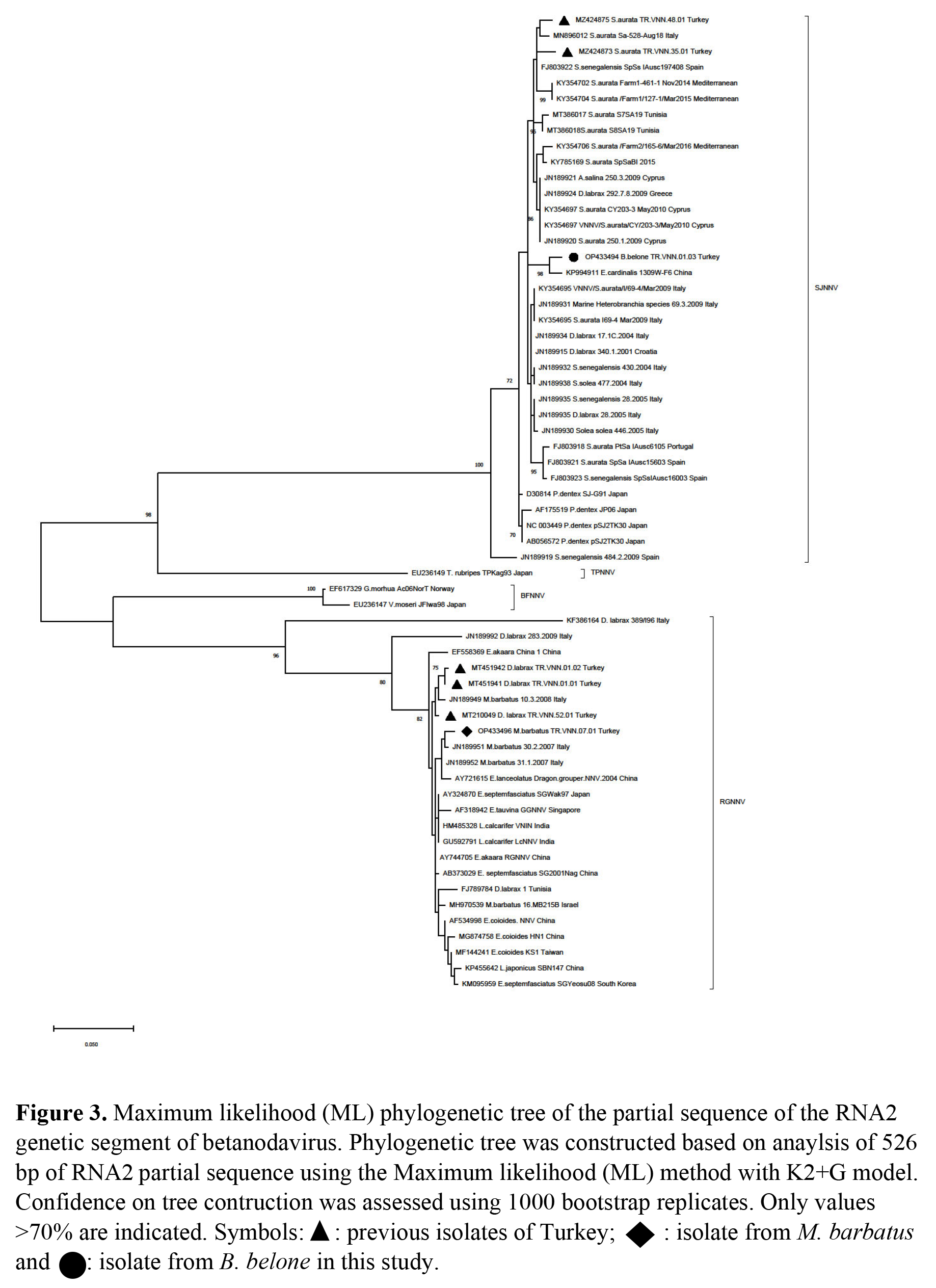
Partial sequencing of TR.VNN.01.03 and TR.VNN.07.01 were performed based on both RNA1 and RNA2 segments. The RNA1 segment of TR.VNN.01.03 and TR.VNN.07.01 were sequenced as 630 and 942 nt, respectively. The RNA2 segment of TR.VNN.01.03 and TR.VNN.07.01 were sequenced as 594 and 526 nt, respectively. Obtained sequences were uploaded to GenBank, and OP433493, OP433495 (RNA1), and OP433494, OP433496 (RNA2) accession numbers were obtained for TR.VNN.01.03 and TR.VNN.07.01, respectively. Isolates of this study were compared with the sequences in GenBank. The RGNNV was determined to be genotype of both isolates according to the RNA1 segment (Figure 2). However, TR.VNN.01.03 was a SJNNV, and TR.VNN.07.01 was a RGNNV genotype according to RNA2 segment (Figure 3). Thus, the TR.VNN.01.03 isolate was reassortant RGNNV/SJNNV, whereas the TR.VNN.07.01 isolate was RGNNV genotype. Interestingly, each of the isolates were detected in different location (Iskenderun and Antalya bays), from different fish species (garfish and red mullet fish), and belonged to the different genotypes (RGNNV/SJNNV and RGNNV). A 610 bp product of the RNA1 segment and 526 bp of RNA2 segment were used to perform further comparisons with other isolates in phylogenetic analysis.


In the phylogenetic analysis of RNA1 segment of TR.VNN.07.01, nt similarity was 98.22% to Italy isolates, while their aa proximity was 100%. TR.VNN.07.01 was found to be 94-95% similar to Turkish isolates previously detected in the Mediterranean (TR.VNN.01.01, TR.VNN.01.02) (Kaplan and Karaoğlu 2021), but aa proximity was 99.47%. The RNA2 segment of TR.VNN.07.01 isolate was 99.22% and 99.02% similar to Italy and China isolates, respectively. The aa proximity was 100% similar to many isolates from both the Mediterranean and the China, India, and Japan coasts (Appendix 1). NNVs previously isolated from red mullet fish worldwide (Berzak et al. 2019; Bitchava et al. 2019; Ciulli et al. 2007; Panzarin et al. 2012) were also examined only in proximity-similarity among themselves. However, because the gene regions where previous researcher’s studied were different, an evaluation could be made on a very short gene region isolated in İtaly, Greece and Israel. According to RNA1 (464 bp) and RNA2 (309 bp) nt similarity, the closest isolate was 31.1.2007 isolated in Italy with 98.46% (aa; 100%) and 99.01% (aa; 100%), respectively (data not shown).
Phylogenetic analysis of RNA1 segment of TR.VNN.01.03 revealed the closest relatedness was Portugal isolates with 97.85 % nt similarity. Furthermore, there was 97.87% similarity in aa with many isolates isolated from the Mediterranean Sea by Turkey, Italy and Portugal. According to the TR.VNN.01.03 isolate’s RNA2 segment, the closest isolate was KP994911 isolated from E. cardinalis in China with 98.46% similarity, the proximity of aa was 98.85% with the same isolate, and the similarity was 97.1% with the majority of Mediterranean isolates used in the phylogenetic study (Appendix-1). Around the world, only in the Mediterranean Sea was NNV reported in one garfish (Ciulli et al. 2007). However, since there were no records in the studies conducted in GenBank, we could not make a comparison with the NNV isolated from the garfish we found in the present study.
Discussion
Study of wild fish in VNN endemic regions in Turkey resulted in finding the virus presence in garfish and red mullet. While the RGNNV/SJNNV reassortant strain was detected in garfish, the RGNNV genotype was detected in red mullet. Both NNV positive fish were asymptomatic and viral loads (>35 Ct) were low in garfish, as expected; but the viral loads in red mullets were very high (22-25 Ct). This suggests that red mullet may play a more significant role in the transmission of NNV. A study of farmed and wild thicklip grey mullets supported the alternative theory. Researchers detected NNV genome in mullets after confirmation of outbreaks in sea bass (Zrnčić et al. 2022). However, much more data and validation are necessary to prove this hypothesis, farm to wild or wild to farm.
VNN is endemic in the Mediterranean basin, and the disease has occurred many times in both farm and wild fish (Berzak et al. 2019; Costa and Thompson 2016; Munday, Kwang, and Moody 2002; OIE 2017; Toffan et al. 2017). RGNNV is the most common genotype in this region (Costa and Thompson 2016), but the SJNNV genotype was also present in the Iberian Peninsula in 2009 (Panzarin et al. 2012). Reassortant RGNNV/SJNNV and SJNNV/RGNNV strains have only been reported in the Mediterranean Sea (He and Teng 2015). NNV has been detected clinically and subclinically in sea bass in Turkey; however, it has only been detected subclinically in sea bream thus far. The first NNV was first detected in Turkey in clinically infected sea bass on the Mediterranean coast in 2011 (Özkan Özyer et al. 2014). In ongoing studies, NNV has been detected frequently in sea bass and the RGNNV genotype was identified (Kalaycı et al. 2016; Kaplan et al. 2021; Kaplan and Karaoğlu 2021). The RGNNV/SJNNV genotype was also isolated from asymptomatic sea bream only in the Aegean Sea (Kaplan, Pekmez, Cagirgan, et al. 2022).
NNV research on wildlife along the Turkish Mediterranean coast has not been documented. So, in terms of taking epidemiological pictures, this research is essential. Here, multiple sampling was performed at five different points in different fish species, and as a result, both RGNNV (red mullet) and RGNNV/SJNNV (garfish) genotypes were isolated. The RGNNV/SJNNV genotype was detected for the first time on the Mediterranean coast of Turkey. Despite the fact that the RGNNV/SJNNV genotype was found in garfish (Ciulli et al. 2007), it was not possible for molecular comparisons to be performed since the records for this genotype were not accessible in GenBank. However, sampling had been performed many times in farms in the region where the garfish were collected (Kaplan and Karaoğlu 2021) and only the RGNNV genotype (TR.VNN.01.01 and TR.VNN.01.02) was identified. Two viruses isolated from the same location, TR.VNN.01.03 and TR.VNN.01.01, show a difference of 5% in the RNA1 segment, suggesting that either different viruses circulate in this region or mutations may have occurred in these segments. However, to clarify this, it is necessary to conduct whole genome studies of the viruses and subsequently molecular evolution studies. Here, the detection of the RGNNV/SJNNV genotype was surprising. In molecular analysis, TR.VNN.01.03 isolated from garfish was closer to the Portuguese reassortants according to the RNA1 segment, while it was closer to the Chinese isolates than the Mediterranean isolates according to the RNA2 segment. As previously reported in the literature, betanodavirus can transmit between different species independently of the host (Costa and Thompson 2016). More importantly, it suggests that different genotypes can circulate across different geographic regions. As a result, there is a constant flow to the Mediterranean Sea through the Red Sea, and the species exotic to the Mediterranean Sea come here from the Indian Ocean through the Red Sea (Al-Sulaimani et al. 2022; Mehanna and Osman 2022; Tomanić et al. 2022). This fact supports the idea that exotic virus species can be transferred through exotic fish by different geographical regions. Additionally, the RGNNV genotype suggests that high diversity among genetic clusters may play a critical role in virus evolution (Sethi et al. 2018). Epistasis and mutation resulting from reassortants within RGNNV may also shape the evolutionary process and diversity of RGNNV (Bitchava et al. 2019). Considering the genome characteristics of betanodavirus, both the segmented structure and especially the RNA1 segment being more prone to mutation, combined with the migration of viruses or exotic fish species, may lead to even more negative consequences. In addition, global warming causes significant changes in aquatic environmental parameters, which may affect the epidemiology of aquatic animal diseases as in other animal diseases (Bandín and Souto 2020). Worse still, this may exacerbate the disease severity in aquaculture worldwide (Bitchava et al. 2019). Rapid increases in viral load due to high numbers and temperature changes in wild asymptomatic species infected with NNV, particularly in Asian and Mediterranean countries, may cause an increase in deaths (Bandín and Souto 2020; Souto, Olveira, and Bandín 2015). In addition, increasing ocean temperatures may increase the number of reported cases of wild species and cause the decline of some endangered species (Bandín and Souto 2020). VNN outbreaks occur mostly in warm conditions (Doan et al. 2016). It is also possible for wild fish species to transmit the virus to farms or vice versa (Bitchava et al. 2019; Zrnčić et al. 2022).
In the molecular analysis of the TR.VNN.07.01 isolated from red mullet, the fact that it is similar to the RGNNV isolates previously isolated from the Mediterranean according to both RNA1 and RNA2 segments, shows that similar virus circulation in red mullets continues in a wide area in this region. Considering that red mullet fish inhabit deeper regions, both the high positivity rate (25/50) and the high viral loads found show that red mullets can play a crucial role in the epidemiology of betanodavirus. With this study, NNV isolation from red mullets has been performed for the first time on the Mediterranean coast of Turkey. The high positivity rate poses a risk in this region. Considering aquaculture in Turkey, further studies on betanodavirus in red mullets in both Mediterranean and Aegean seas can contribute to sustainable cultural fisheries.
NNV has been detected in more than 120 wild aquatic species globally (Bandín and Souto 2020). Although most are asymptomatic, NNV-related mortality has been reported in milk fish (Chanos chanos) (Sethi et al. 2018), European sea bass (Vendramin et al. 2013), mullet (Liza aurata and L. saliens) (Zorriehzahra et al. 2016) and wild grouper (E. costae and E. marginatus) (Kara et al. 2014; Nishioka et al. 2016; Valencia et al. 2019; Vendramin et al. 2013). In previous studies in the Mediterranean, NNV were detected in asymptomatic (Berzak et al. 2019; Bovo et al. 2011; Ciulli et al. 2007; Panzarin et al. 2012) or clinically infected (Haddad-Boubaker et al. 2014; Lopez-Jimena et al. 2010; Maltese et al. 2005; Terlizzi, Tedesco, and Patarnello 2012; Vendramin et al. 2013; Valencia et al. 2019) wild fish many times. Although RGNNV was identified frequently in wild populations in the Mediterranean (Bandín and Souto 2020), RGNNV/SJNNV reassortant was isolated from wild Mediterranean horse mackerel (Trachurus mediterraneus) and greater amberjack (Seriola dumerili) in Greece (Bitchava et al. 2019), thicklip grey mullet (Chelon labrosus) in Spain (Haddad-Boubaker et al. 2014). In Asia, the RGNNV/SJNNV was isolated from whitefin trevally (Carangoides equula) and Japanese jack mackerel (T. japonicus) species (Gomez et al. 2004; Liu et al. 2014). In this study, an RGNNV/SJNNV reassortant was identified in garfish in the Mediterranean. With new research, the number of fish species involved in the spread of the betanodavirus is increasing day by day. The increase in risk factors constantly reminds us of the increase in biosecurity measures on farms and their environments. However, in the future, multivalent vaccines may be necessary, rather than monovalent vaccines to prevent outbreaks. On the other hand, increasing fish species in betanodavirus epidemiology poses a threat to aquaculture, which shows the importance of screening studies on wildlife or farms. Asymptomatic infected fish are eaten by predator (hunter‒predator) fish and get contaminated with infected stools of these fish. Another possible transmission of the disease is the feeding of broodstocks with raw fish (Mori et al. 2005). For example, after detecting NNV in wildlife, it could be detected in a farm in a short time (Berzak et al. 2019). Therefore, research on wild fish is critical in sustaining the future of aquaculture industries. The close viral sequences between betanodavirus isolates from wild fish and those isolated during clinical disease outbreaks in cultured fish in the same region in previous years show permanent circulation and interaction and transmission between wildlife and farm fish (Vendramin et al. 2013). Horizontal transmission among subclinical fish after migration or commercial importation for cultural fishery production can play a significant role in new outbreaks (Valencia et al. 2019). Information on the health status of specific wild fish populations is also crucial for aquaculture (Moreno et al. 2014). Considering the origins of NNV isolates detected thus far in Turkey, although most were asymptomatic, high aquaculture production capacity in Turkey necessitates early warning systems. To date, studies on NNV screening in Aegean, Mediterranean, or Black Sea regions have been at the farm level (Kaplan et al. 2021; Kaplan and Karaoğlu 2021; Kaplan, Pekmez, Çagirgan, et al. 2022). Additionally, NNV studies were conducted on mollusks in North Aegean and Marmara seas, but no virus was detected (Kaplan, Pekmez, Çagirgan, et al. 2022). However, a screening study involving wild fish in the Mediterranean Sea, where the virus is endemic, most abundant, and circulation is highest has yet to be conducted. This study involved the collection of data on the epidemiology of betanodavirus for the first time in wildlife of this region.
In a conclusion, after evaluating the genetic similarities between isolates in this study and other isolates, the isolates were similar to those found in Italy and the far-east rather than previous Turkish isolates. The first task at this point is to perform NNV screening in wild fish in a much wider area, i.e. in order to obtain whole genome sequences of possible isolates, and follow-up with a phylogenetic study. In the context of sustainable aquaculture production, it will be vital to reveal whole genome sequences of all isolates and to determine the prevalence or differences to be considered for potential vaccines.
Acknowledgments
This study was supported and officially permissed by the Ministry of Agriculture and Forestry of the Republic of Türkiye.